 |
PDBsum entry 2hob

|
|
|
|
|
|

|

|

|

|
|

|
|

|
|

|
|

|
|
|
 |

|

|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Viral protein
|
PDB id
|
|
|
|
2hob
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|

|

|

|
|
|
|
|
|
|
|
 |
Contents |
 |
|
|
|
|
|
|
|
|
|
|
|
|
* Residue conservation analysis
|
|
|
|
|
|
|
|
|
|
|
Enzyme class 2:
|
|
E.C.2.1.1.-
- ?????
|
|
|
|
|
|
|
Enzyme class 3:
|
|
E.C.2.1.1.56
- mRNA (guanine-N(7))-methyltransferase.
|
|
|
|
|
|
|
Reaction:
|
|
a 5'-end (5'-triphosphoguanosine)-ribonucleoside in mRNA + S-adenosyl-L- methionine = a 5'-end (N(7)-methyl 5'-triphosphoguanosine)-ribonucleoside in mRNA + S-adenosyl-L-homocysteine
|
|
|
|
|
|
5'-end (5'-triphosphoguanosine)-ribonucleoside in mRNA
|
+
|
 S-adenosyl-L- methionine
S-adenosyl-L- methionine
|
=
|
5'-end (N(7)-methyl 5'-triphosphoguanosine)-ribonucleoside in mRNA
|
+
|
 S-adenosyl-L-homocysteine
S-adenosyl-L-homocysteine
|
|
|
|
|
|
|
|
|
|
Enzyme class 4:
|
|
E.C.2.1.1.57
- methyltransferase cap1.
|
|
|
|
|
|
|
Reaction:
|
|
a 5'-end (N(7)-methyl 5'-triphosphoguanosine)-ribonucleoside in mRNA + S-adenosyl-L-methionine = a 5'-end (N(7)-methyl 5'-triphosphoguanosine)- (2'-O-methyl-ribonucleoside) in mRNA + S-adenosyl-L-homocysteine + H+
|
|
|
|
|
|
5'-end (N(7)-methyl 5'-triphosphoguanosine)-ribonucleoside in mRNA
|
+
|
S-adenosyl-L-methionine
|
=
|
5'-end (N(7)-methyl 5'-triphosphoguanosine)- (2'-O-methyl-ribonucleoside) in mRNA
|
+
|
S-adenosyl-L-homocysteine
|
+
|
H(+)
|
|
|
|
|
|
|
|
|
|
Enzyme class 5:
|
|
E.C.2.7.7.48
- RNA-directed Rna polymerase.
|
|
|
|
|
|
|
Reaction:
|
|
RNA(n) + a ribonucleoside 5'-triphosphate = RNA(n+1) + diphosphate
|
|
|
|
|
|
 RNA(n)
RNA(n)
|
+
|
ribonucleoside 5'-triphosphate
|
=
|
RNA(n+1)
|
+
|
 diphosphate
diphosphate
|
|
|
|
|
|
|
|
|
|
Enzyme class 6:
|
|
E.C.2.7.7.50
- mRNA guanylyltransferase.
|
|
|
|
|
|
|
Reaction:
|
|
a 5'-end diphospho-ribonucleoside in mRNA + GTP + H+ = a 5'-end (5'-triphosphoguanosine)-ribonucleoside in mRNA + diphosphate
|
|
|
|
|
|
5'-end diphospho-ribonucleoside in mRNA
|
+
|
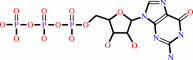 GTP
GTP
|
+
|
H(+)
|
=
|
5'-end (5'-triphosphoguanosine)-ribonucleoside in mRNA
|
+
|
diphosphate
|
|
|
|
|
|
|
|
|
|
Enzyme class 7:
|
|
E.C.3.1.13.-
- ?????
|
|
|
|
|
|
|
Enzyme class 8:
|
|
E.C.3.4.19.12
- ubiquitinyl hydrolase 1.
|
|
|
|
|
|
|
Reaction:
|
|
Thiol-dependent hydrolysis of ester, thiolester, amide, peptide and isopeptide bonds formed by the C-terminal Gly of ubiquitin (a 76-residue protein attached to proteins as an intracellular targeting signal).
|
|
|
|
|
|
Enzyme class 9:
|
|
E.C.3.4.22.-
- ?????
|
|
|
|
|
|
|
Enzyme class 10:
|
|
E.C.3.4.22.69
- Sars coronavirus main proteinase.
|
|
|
|
|
|
|
Enzyme class 11:
|
|
E.C.3.6.4.12
- Dna helicase.
|
|
|
|
|
|
|
Reaction:
|
|
ATP + H2O = ADP + phosphate + H+
|
|
|
|
|
|
 ATP
ATP
|
+
|
H2O
|
=
|
 ADP
ADP
|
+
|
 phosphate
phosphate
|
+
|
H(+)
|
|
|
|
|
|
|
|
|
|
Enzyme class 12:
|
|
E.C.3.6.4.13
- Rna helicase.
|
|
|
|
|
|
|
Reaction:
|
|
ATP + H2O = ADP + phosphate + H+
|
|
|
|
|
|
ATP
|
+
|
H2O
|
=
|
ADP
|
+
|
phosphate
|
+
|
H(+)
|
|
|
|
|
|
|
|
|
|
Enzyme class 13:
|
|
E.C.4.6.1.-
- ?????
|
|
|
|
|
|
|
|
|
|
Note, where more than one E.C. class is given (as above), each may
correspond to a different protein domain or, in the case of polyprotein
precursors, to a different mature protein.
|
|
|
|
Molecule diagrams generated from .mol files obtained from the
KEGG ftp site
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
| |
|
|
| |
|
DOI no:
|
J Mol Biol
366:965-975
(2007)
|
|
PubMed id:
|
|
|
|
|
|
| |
|
Production of authentic SARS-CoV M(pro) with enhanced activity: application as a novel tag-cleavage endopeptidase for protein overproduction.
|
|
X.Xue,
H.Yang,
W.Shen,
Q.Zhao,
J.Li,
K.Yang,
C.Chen,
Y.Jin,
M.Bartlam,
Z.Rao.
|
|
|
|
|
| |
ABSTRACT
|
|
|
|
| |
|
|
The viral proteases have proven to be the most selective and useful for removing
the fusion tags in fusion protein expression systems. As a key enzyme in the
viral life-cycle, the main protease (M(pro)) is most attractive for drug design
targeting the SARS coronavirus (SARS-CoV), the etiological agent responsible for
the outbreak of severe acute respiratory syndrome (SARS) in 2003. In this study,
SARS-CoV M(pro) was used to specifically remove the GST tag in a new fusion
protein expression system. We report a new method to produce wild-type (WT)
SARS-CoV M(pro) with authentic N and C termini, and compare the activity of WT
protease with those of three different types of SARS-CoV M(pro) with additional
residues at the N or C terminus. Our results show that additional residues at
the N terminus, but not at the C terminus, of M(pro) are detrimental to enzyme
activity. To explain this, the crystal structures of WT SARS-CoV M(pro) and its
complex with a Michael acceptor inhibitor were determined to 1.6 Angstroms and
1.95 Angstroms resolution respectively. These crystal structures reveal that the
first residue of this protease is important for sustaining the substrate-binding
pocket and inhibitor binding. This study suggests that SARS-CoV M(pro) could
serve as a new tag-cleavage endopeptidase for protein overproduction, and the WT
SARS-CoV M(pro) is more appropriate for mechanistic characterization and
inhibitor design.
|
|
|
|
|
|
| |
Selected figure(s)
|
|
|
|
| |
|
|

|
|

|
|
Figure 3.
Figure 3. Superposition of the S1 pockets of GPLGS-WT and WT
SARS-CoV M^pro (in stereo). (a) Superposition of the S1 pockets
in protomer A of GPLGS-WT and that of protomer A* of WT SARS-CoV
M^pro. Protomer A* of WT is in blue; protomer A of GPLGS-WT is
in yellow; protomer B* of WT is in magenta; protomer B of
GPLGS-WT is in red. In the WT structure, the amino group (NH[2])
of Ser1 in protomer B* donates a 3.0 Å hydrogen bond to
the carboxylate group of Glu166 and a 2.7 Å hydrogen bond
to the main-chain carbonyl group of Phe140 in protomer A*,
stabilizing the S1 pocket. The NH of Gly143 moves 0.8 Å
toward the activity site; the main chain of residues 142-143
moves toward the S1 subsite; the side-chain of Asn-A*142 flips
over with a 6 Å shift compared with protomer A of
GPLGS-WT. (b) Superposition of the S1 pockets in protomer B of
GPLGS-WT and that of Protomer A* of WT SARS-CoV M^pro. Protomer
A* of WT is in blue; protomer B of GPLGS-WT is in yellow;
protomer B* of WT is in magenta; protomer A of GPLGS-WT is in
red. The S1 pocket of protomer B collapses partly with
reorientation of Glu166 and residues 140–143. No electron
density was visible for residues A1 and A2. Figure 3.
Superposition of the S1 pockets of GPLGS-WT and WT SARS-CoV
M^pro (in stereo). (a) Superposition of the S1 pockets in
protomer A of GPLGS-WT and that of protomer A* of WT SARS-CoV
M^pro. Protomer A* of WT is in blue; protomer A of GPLGS-WT is
in yellow; protomer B* of WT is in magenta; protomer B of
GPLGS-WT is in red. In the WT structure, the amino group (NH[2])
of Ser1 in protomer B* donates a 3.0 Å hydrogen bond to
the carboxylate group of Glu166 and a 2.7 Å hydrogen bond
to the main-chain carbonyl group of Phe140 in protomer A*,
stabilizing the S1 pocket. The NH of Gly143 moves 0.8 Å
toward the activity site; the main chain of residues 142-143
moves toward the S1 subsite; the side-chain of Asn-A*142 flips
over with a 6 Å shift compared with protomer A of
GPLGS-WT. (b) Superposition of the S1 pockets in protomer B of
GPLGS-WT and that of Protomer A* of WT SARS-CoV M^pro. Protomer
A* of WT is in blue; protomer B of GPLGS-WT is in yellow;
protomer B* of WT is in magenta; protomer A of GPLGS-WT is in
red. The S1 pocket of protomer B collapses partly with
reorientation of Glu166 and residues 140–143. No electron
density was visible for residues A1 and A2.
|
|
Figure 4.
Figure 4. Differences between the complex structures of WT and
GPLGS-WT. (a) Inhibitor N3. (b) Superposition of the
substrate-binding pockets in protomer A of GPLGS-WT and that in
protomer A* of WT. In the WT-N3 complex structure, the NH[2]
group of Ser1 in protomer B* was still hydrogen-bonded to the
carboxylate group of Glu166 and the carbonyl group of Phe140 in
protomer A*, stabilizing the S1 pocket. In the GPLGS-WT-N3
complex structure, however, the two hydrogen bonds described
above were not found. Instead, an ordered water molecule was
observed in the S1 pocket. Protomer A* of WT is in blue;
protomer A of GPLGS-WT is in yellow; inhibitor N3 (complexed
with WT) is in magenta; inhibitor N3 (complexed with GPLGS-WT)
is in red; protomer B* of WT is in green; protomer B of GPLGS-WT
is in cyan. Figure 4. Differences between the complex
structures of WT and GPLGS-WT. (a) Inhibitor N3. (b)
Superposition of the substrate-binding pockets in protomer A of
GPLGS-WT and that in protomer A* of WT. In the WT-N3 complex
structure, the NH[2] group of Ser1 in protomer B* was still
hydrogen-bonded to the carboxylate group of Glu166 and the
carbonyl group of Phe140 in protomer A*, stabilizing the S1
pocket. In the GPLGS-WT-N3 complex structure, however, the two
hydrogen bonds described above were not found. Instead, an
ordered water molecule was observed in the S1 pocket. Protomer
A* of WT is in blue; protomer A of GPLGS-WT is in yellow;
inhibitor N3 (complexed with WT) is in magenta; inhibitor N3
(complexed with GPLGS-WT) is in red; protomer B* of WT is in
green; protomer B of GPLGS-WT is in cyan.
|
|
|
|
|
|
| |
The above figures are
reprinted
by permission from Elsevier:
J Mol Biol
(2007,
366,
965-975)
copyright 2007.
|
|
| |
Figures were
selected
by an automated process.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
 |
 |
|
Literature references that cite this PDB file's key reference
|
|
|
| |
PubMed id
|
|
Reference
|
|
|
|
|
|
X.Jia,
R.Singh,
S.Homann,
H.Yang,
J.Guatelli,
and
Y.Xiong
(2012).
Structural basis of evasion of cellular adaptive immunity by HIV-1 Nef.
|
| |
Nat Struct Mol Biol,
19,
701-706.
|
|
|
PDB codes:
|
|
|
|
|
|
|
|
R.N.Kostoff
(2010).
The highly cited SARS research literature.
|
| |
Crit Rev Microbiol,
36,
299-317.
|
|
|
|
|
|
|
A.K.Ghosh,
G.Gong,
V.Grum-Tokars,
D.C.Mulhearn,
S.C.Baker,
M.Coughlin,
B.S.Prabhakar,
K.Sleeman,
M.E.Johnson,
and
A.D.Mesecar
(2008).
Design, synthesis and antiviral efficacy of a series of potent chloropyridyl ester-derived SARS-CoV 3CLpro inhibitors.
|
| |
Bioorg Med Chem Lett,
18,
5684-5688.
|
|
|
|
|
|
|
J.S.Sparks,
E.F.Donaldson,
X.Lu,
R.S.Baric,
and
M.R.Denison
(2008).
A novel mutation in murine hepatitis virus nsp5, the viral 3C-like proteinase, causes temperature-sensitive defects in viral growth and protein processing.
|
| |
J Virol,
82,
5999-6008.
|
|
|
|
|
|
|
J.Shi,
J.Sivaraman,
and
J.Song
(2008).
Mechanism for controlling the dimer-monomer switch and coupling dimerization to catalysis of the severe acute respiratory syndrome coronavirus 3C-like protease.
|
| |
J Virol,
82,
4620-4629.
|
|
|
PDB code:
|
|
|
|
|
|
|
|
N.Zhong,
S.Zhang,
P.Zou,
J.Chen,
X.Kang,
Z.Li,
C.Liang,
C.Jin,
and
B.Xia
(2008).
Without its N-finger, the main protease of severe acute respiratory syndrome coronavirus can form a novel dimer through its C-terminal domain.
|
| |
J Virol,
82,
4227-4234.
|
|
|
|
|
|
|
S.Chen,
T.Hu,
J.Zhang,
J.Chen,
K.Chen,
J.Ding,
H.Jiang,
and
X.Shen
(2008).
Mutation of Gly-11 on the dimer interface results in the complete crystallographic dimer dissociation of severe acute respiratory syndrome coronavirus 3C-like protease: crystal structure with molecular dynamics simulations.
|
| |
J Biol Chem,
283,
554-564.
|
|
|
PDB code:
|
|
|
|
|
|
|
|
X.Xue,
H.Yu,
H.Yang,
F.Xue,
Z.Wu,
W.Shen,
J.Li,
Z.Zhou,
Y.Ding,
Q.Zhao,
X.C.Zhang,
M.Liao,
M.Bartlam,
and
Z.Rao
(2008).
Structures of two coronavirus main proteases: implications for substrate binding and antiviral drug design.
|
| |
J Virol,
82,
2515-2527.
|
|
|
PDB codes:
|
|
|
|
|
|
|
|
D.Plewczynski,
M.Hoffmann,
M.von Grotthuss,
K.Ginalski,
and
L.Rychewski
(2007).
In silico prediction of SARS protease inhibitors by virtual high throughput screening.
|
| |
Chem Biol Drug Des,
69,
269-279.
|
|
|
|
|
|
The most recent references are shown first.
Citation data come partly from CiteXplore and partly
from an automated harvesting procedure. Note that this is likely to be
only a partial list as not all journals are covered by
either method. However, we are continually building up the citation data
so more and more references will be included with time.
Where a reference describes a PDB structure, the PDB
codes are
shown on the right.
|
|


 Links
Links







































