 |
PDBsum entry 2fe8

|
|
|
|
|
 |
Contents |
 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
* Residue conservation analysis
|
|
|
|
|
|
|
|
|
|
|
Enzyme class 2:
|
|
E.C.2.1.1.-
- ?????
|
|
|
|
|
|
|
Enzyme class 3:
|
|
E.C.2.1.1.56
- mRNA (guanine-N(7))-methyltransferase.
|
|
|
|
|
|
|
Reaction:
|
|
a 5'-end (5'-triphosphoguanosine)-ribonucleoside in mRNA + S-adenosyl-L- methionine = a 5'-end (N(7)-methyl 5'-triphosphoguanosine)-ribonucleoside in mRNA + S-adenosyl-L-homocysteine
|
|
|
|
|
|
5'-end (5'-triphosphoguanosine)-ribonucleoside in mRNA
|
+
|
 S-adenosyl-L- methionine
S-adenosyl-L- methionine
|
=
|
5'-end (N(7)-methyl 5'-triphosphoguanosine)-ribonucleoside in mRNA
|
+
|
 S-adenosyl-L-homocysteine
S-adenosyl-L-homocysteine
|
|
|
|
|
|
|
|
|
|
Enzyme class 4:
|
|
E.C.2.1.1.57
- methyltransferase cap1.
|
|
|
|
|
|
|
Reaction:
|
|
a 5'-end (N(7)-methyl 5'-triphosphoguanosine)-ribonucleoside in mRNA + S-adenosyl-L-methionine = a 5'-end (N(7)-methyl 5'-triphosphoguanosine)- (2'-O-methyl-ribonucleoside) in mRNA + S-adenosyl-L-homocysteine + H+
|
|
|
|
|
|
5'-end (N(7)-methyl 5'-triphosphoguanosine)-ribonucleoside in mRNA
|
+
|
S-adenosyl-L-methionine
|
=
|
5'-end (N(7)-methyl 5'-triphosphoguanosine)- (2'-O-methyl-ribonucleoside) in mRNA
|
+
|
S-adenosyl-L-homocysteine
|
+
|
H(+)
|
|
|
|
|
|
|
|
|
|
Enzyme class 5:
|
|
E.C.2.7.7.48
- RNA-directed Rna polymerase.
|
|
|
|
|
|
|
Reaction:
|
|
RNA(n) + a ribonucleoside 5'-triphosphate = RNA(n+1) + diphosphate
|
|
|
|
|
|
 RNA(n)
RNA(n)
|
+
|
ribonucleoside 5'-triphosphate
|
=
|
RNA(n+1)
|
+
|
 diphosphate
diphosphate
|
|
|
|
|
|
|
|
|
|
Enzyme class 6:
|
|
E.C.2.7.7.50
- mRNA guanylyltransferase.
|
|
|
|
|
|
|
Reaction:
|
|
a 5'-end diphospho-ribonucleoside in mRNA + GTP + H+ = a 5'-end (5'-triphosphoguanosine)-ribonucleoside in mRNA + diphosphate
|
|
|
|
|
|
5'-end diphospho-ribonucleoside in mRNA
|
+
|
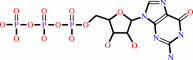 GTP
GTP
|
+
|
H(+)
|
=
|
5'-end (5'-triphosphoguanosine)-ribonucleoside in mRNA
|
+
|
diphosphate
|
|
|
|
|
|
|
|
|
|
Enzyme class 7:
|
|
E.C.3.1.13.-
- ?????
|
|
|
|
|
|
|
Enzyme class 8:
|
|
E.C.3.4.19.12
- ubiquitinyl hydrolase 1.
|
|
|
|
|
|
|
Reaction:
|
|
Thiol-dependent hydrolysis of ester, thiolester, amide, peptide and isopeptide bonds formed by the C-terminal Gly of ubiquitin (a 76-residue protein attached to proteins as an intracellular targeting signal).
|
|
|
|
|
|
Enzyme class 9:
|
|
E.C.3.4.22.-
- ?????
|
|
|
|
|
|
|
Enzyme class 10:
|
|
E.C.3.4.22.69
- Sars coronavirus main proteinase.
|
|
|
|
|
|
|
Enzyme class 11:
|
|
E.C.3.6.4.12
- Dna helicase.
|
|
|
|
|
|
|
Reaction:
|
|
ATP + H2O = ADP + phosphate + H+
|
|
|
|
|
|
 ATP
ATP
|
+
|
H2O
|
=
|
 ADP
ADP
|
+
|
 phosphate
phosphate
|
+
|
H(+)
|
|
|
|
|
|
|
|
|
|
Enzyme class 12:
|
|
E.C.3.6.4.13
- Rna helicase.
|
|
|
|
|
|
|
Reaction:
|
|
ATP + H2O = ADP + phosphate + H+
|
|
|
|
|
|
ATP
|
+
|
H2O
|
=
|
ADP
|
+
|
phosphate
|
+
|
H(+)
|
|
|
|
|
|
|
|
|
|
Enzyme class 13:
|
|
E.C.4.6.1.-
- ?????
|
|
|
|
|
|
|
|
|
|
Note, where more than one E.C. class is given (as above), each may
correspond to a different protein domain or, in the case of polyprotein
precursors, to a different mature protein.
|
|
|
|
Molecule diagrams generated from .mol files obtained from the
KEGG ftp site
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
| |
|
|
| |
|
DOI no:
|
Proc Natl Acad Sci U S A
103:5717-5722
(2006)
|
|
PubMed id:
|
|
|
|
|
|
| |
|
Severe acute respiratory syndrome coronavirus papain-like protease: structure of a viral deubiquitinating enzyme.
|
|
K.Ratia,
K.S.Saikatendu,
B.D.Santarsiero,
N.Barretto,
S.C.Baker,
R.C.Stevens,
A.D.Mesecar.
|
|
|
|
|
| |
ABSTRACT
|
|
|
|
| |
|
|
Replication of severe acute respiratory syndrome (SARS) coronavirus (SARS-CoV)
requires proteolytic processing of the replicase polyprotein by two viral
cysteine proteases, a chymotrypsin-like protease (3CLpro) and a papain-like
protease (PLpro). These proteases are important targets for development of
antiviral drugs that would inhibit viral replication and reduce mortality
associated with outbreaks of SARS-CoV. In this work, we describe the 1.85-A
crystal structure of the catalytic core of SARS-CoV PLpro and show that the
overall architecture adopts a fold closely resembling that of known
deubiquitinating enzymes. Key features, however, distinguish PLpro from
characterized deubiquitinating enzymes, including an intact zinc-binding motif,
an unobstructed catalytically competent active site, and the presence of an
intriguing, ubiquitin-like N-terminal domain. To gain insight into the
active-site recognition of the C-terminal tail of ubiquitin and the related LXGG
motif, we propose a model of PLpro in complex with ubiquitin-aldehyde that
reveals well defined sites within the catalytic cleft that help to account for
strict substrate-recognition motifs.
|
|
|
|
|
|
| |
Selected figure(s)
|
|
|
|
| |
|
|

|
|

|
|
Figure 4.
Fig. 4. The SARS-CoV PLpro and USP14 active sites. (A)
SARS-CoV PLpro catalytic triad residues, C112, H273, and D287,
and other important active-site residues. Distances between
residues are indicated in angstroms. The hydrogen bond between
D109 and W97 is indicated by an arrow. (B) Comparison of USP14
and SARS-CoV PLpro BL1 and BL2 loop regions. Corresponding
regions of unbound USP14 (red), Ubal-complexed USP14 (yellow),
and PLpro (blue) are shown superimposed. The BL1 and BL2 loop
regions are indicated. The BL1 loop region of PLpro is colored
in green. The catalytic triad residues are shown by a
ball-and-stick representation.
|
|
Figure 5.
Fig. 5. Comparison of the ubiquitin-binding surfaces of
HAUSP, USP14, and PLpro based on modeling studies. For modeling
ubiquitin into the SARS-CoV PLpro active site, the structures of
the Ubal-bound forms of HAUSP (1nbf) and USP14 (2ayo) were
superimposed onto the PLpro structure and analyzed. Contacts at
the C-terminal tail of ubiquitin and two interacting surfaces of
the PLpro palm domain were manually edited and minimized by
using CNS. The ubiquitin molecule is shown as a ribbon diagram.
|
|
|
|
| |
Figures were
selected
by an automated process.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
 |
 |
|
Literature references that cite this PDB file's key reference
|
|
|
| |
PubMed id
|
|
Reference
|
|
|
|
|
|
S.W.Li,
C.C.Lai,
J.F.Ping,
F.J.Tsai,
L.Wan,
Y.J.Lin,
S.H.Kung,
and
C.W.Lin
(2011).
Severe acute respiratory syndrome coronavirus papain-like protease suppressed alpha interferon-induced responses through downregulation of extracellular signal-regulated kinase 1-mediated signalling pathways.
|
| |
J Gen Virol,
92,
1127-1140.
|
|
|
|
|
|
|
J.A.Wojdyla,
I.Manolaridis,
P.B.van Kasteren,
M.Kikkert,
E.J.Snijder,
A.E.Gorbalenya,
and
P.A.Tucker
(2010).
Papain-like protease 1 from transmissible gastroenteritis virus: crystal structure and enzymatic activity toward viral and cellular substrates.
|
| |
J Virol,
84,
10063-10073.
|
|
|
PDB code:
|
|
|
|
|
|
|
|
K.R.Hurst,
R.Ye,
S.J.Goebel,
P.Jayaraman,
and
P.S.Masters
(2010).
An interaction between the nucleocapsid protein and a component of the replicase-transcriptase complex is crucial for the infectivity of coronavirus genomic RNA.
|
| |
J Virol,
84,
10276-10288.
|
|
|
|
|
|
|
M.A.Clementz,
Z.Chen,
B.S.Banach,
Y.Wang,
L.Sun,
K.Ratia,
Y.M.Baez-Santos,
J.Wang,
J.Takayama,
A.K.Ghosh,
K.Li,
A.D.Mesecar,
and
S.C.Baker
(2010).
Deubiquitinating and interferon antagonism activities of coronavirus papain-like proteases.
|
| |
J Virol,
84,
4619-4629.
|
|
|
|
|
|
|
M.J.Gadlage,
and
M.R.Denison
(2010).
Exchange of the coronavirus replicase polyprotein cleavage sites alters protease specificity and processing.
|
| |
J Virol,
84,
6894-6898.
|
|
|
|
|
|
|
M.Raaben,
C.C.Posthuma,
M.H.Verheije,
E.G.te Lintelo,
M.Kikkert,
J.W.Drijfhout,
E.J.Snijder,
P.J.Rottier,
and
C.A.de Haan
(2010).
The ubiquitin-proteasome system plays an important role during various stages of the coronavirus infection cycle.
|
| |
J Virol,
84,
7869-7879.
|
|
|
|
|
|
|
M.Y.Tsai,
W.H.Chang,
J.Y.Liang,
L.L.Lin,
G.G.Chang,
and
H.P.Chang
(2010).
Essential covalent linkage between the chymotrypsin-like domain and the extra domain of the SARS-CoV main protease.
|
| |
J Biochem,
148,
349-358.
|
|
|
|
|
|
|
R.N.Kostoff
(2010).
The highly cited SARS research literature.
|
| |
Crit Rev Microbiol,
36,
299-317.
|
|
|
|
|
|
|
A.Chatterjee,
M.A.Johnson,
P.Serrano,
B.Pedrini,
J.S.Joseph,
B.W.Neuman,
K.Saikatendu,
M.J.Buchmeier,
P.Kuhn,
and
K.Wüthrich
(2009).
Nuclear magnetic resonance structure shows that the severe acute respiratory syndrome coronavirus-unique domain contains a macrodomain fold.
|
| |
J Virol,
83,
1823-1836.
|
|
|
PDB codes:
|
|
|
|
|
|
|
|
I.Manolaridis,
J.A.Wojdyla,
S.Panjikar,
E.J.Snijder,
A.E.Gorbalenya,
H.Berglind,
P.Nordlund,
B.Coutard,
and
P.A.Tucker
(2009).
Structure of the C-terminal domain of nsp4 from feline coronavirus.
|
| |
Acta Crystallogr D Biol Crystallogr,
65,
839-846.
|
|
|
PDB code:
|
|
|
|
|
|
|
|
J.Han,
M.S.Rutherford,
and
K.S.Faaberg
(2009).
The porcine reproductive and respiratory syndrome virus nsp2 cysteine protease domain possesses both trans- and cis-cleavage activities.
|
| |
J Virol,
83,
9449-9463.
|
|
|
|
|
|
|
J.Tan,
C.Vonrhein,
O.S.Smart,
G.Bricogne,
M.Bollati,
Y.Kusov,
G.Hansen,
J.R.Mesters,
C.L.Schmidt,
and
R.Hilgenfeld
(2009).
The SARS-Unique Domain (SUD) of SARS Coronavirus Contains Two Macrodomains That Bind G-Quadruplexes.
|
| |
PLoS Pathog,
5,
e1000428.
|
|
|
PDB codes:
|
|
|
|
|
|
|
|
M.Frieman,
K.Ratia,
R.E.Johnston,
A.D.Mesecar,
and
R.S.Baric
(2009).
Severe acute respiratory syndrome coronavirus papain-like protease ubiquitin-like domain and catalytic domain regulate antagonism of IRF3 and NF-kappaB signaling.
|
| |
J Virol,
83,
6689-6705.
|
|
|
|
|
|
|
P.Serrano,
M.A.Johnson,
A.Chatterjee,
B.W.Neuman,
J.S.Joseph,
M.J.Buchmeier,
P.Kuhn,
and
K.Wüthrich
(2009).
Nuclear magnetic resonance structure of the nucleic acid-binding domain of severe acute respiratory syndrome coronavirus nonstructural protein 3.
|
| |
J Virol,
83,
12998-13008.
|
|
|
PDB code:
|
|
|
|
|
|
|
|
S.Perlman,
and
J.Netland
(2009).
Coronaviruses post-SARS: update on replication and pathogenesis.
|
| |
Nat Rev Microbiol,
7,
439-450.
|
|
|
|
|
|
|
Y.Zhou,
W.P.Tzeng,
Y.Ye,
Y.Huang,
S.Li,
Y.Chen,
T.K.Frey,
and
J.J.Yang
(2009).
A cysteine-rich metal-binding domain from rubella virus non-structural protein is essential for viral protease activity and virus replication.
|
| |
Biochem J,
417,
477-483.
|
|
|
|
|
|
|
B.Canard,
J.S.Joseph,
and
P.Kuhn
(2008).
International research networks in viral structural proteomics: again, lessons from SARS.
|
| |
Antiviral Res,
78,
47-50.
|
|
|
|
|
|
|
B.W.Neuman,
J.S.Joseph,
K.S.Saikatendu,
P.Serrano,
A.Chatterjee,
M.A.Johnson,
L.Liao,
J.P.Klaus,
J.R.Yates,
K.Wüthrich,
R.C.Stevens,
M.J.Buchmeier,
and
P.Kuhn
(2008).
Proteomics analysis unravels the functional repertoire of coronavirus nonstructural protein 3.
|
| |
J Virol,
82,
5279-5294.
|
|
|
|
|
|
|
C.Zhang,
O.Crasta,
S.Cammer,
R.Will,
R.Kenyon,
D.Sullivan,
Q.Yu,
W.Sun,
R.Jha,
D.Liu,
T.Xue,
Y.Zhang,
M.Moore,
P.McGarvey,
H.Huang,
Y.Chen,
J.Zhang,
R.Mazumder,
C.Wu,
and
B.Sobral
(2008).
An emerging cyberinfrastructure for biodefense pathogen and pathogen-host data.
|
| |
Nucleic Acids Res,
36,
D884-D891.
|
|
|
|
|
|
|
J.Pan,
X.Peng,
Y.Gao,
Z.Li,
X.Lu,
Y.Chen,
M.Ishaq,
D.Liu,
M.L.Dediego,
L.Enjuanes,
and
D.Guo
(2008).
Genome-wide analysis of protein-protein interactions and involvement of viral proteins in SARS-CoV replication.
|
| |
PLoS ONE,
3,
e3299.
|
|
|
|
|
|
|
K.Ratia,
S.Pegan,
J.Takayama,
K.Sleeman,
M.Coughlin,
S.Baliji,
R.Chaudhuri,
W.Fu,
B.S.Prabhakar,
M.E.Johnson,
S.C.Baker,
A.K.Ghosh,
and
A.D.Mesecar
(2008).
A noncovalent class of papain-like protease/deubiquitinase inhibitors blocks SARS virus replication.
|
| |
Proc Natl Acad Sci U S A,
105,
16119-16124.
|
|
|
|
|
|
|
M.Bartlam,
X.Xue,
and
Z.Rao
(2008).
The search for a structural basis for therapeutic intervention against the SARS coronavirus.
|
| |
Acta Crystallogr A,
64,
204-213.
|
|
|
|
|
|
|
M.Drag,
J.Mikolajczyk,
M.Bekes,
F.E.Reyes-Turcu,
J.A.Ellman,
K.D.Wilkinson,
and
G.S.Salvesen
(2008).
Positional-scanning fluorigenic substrate libraries reveal unexpected specificity determinants of DUBs (deubiquitinating enzymes).
|
| |
Biochem J,
415,
367-375.
|
|
|
|
|
|
|
P.Serrano,
M.A.Johnson,
A.Chatterjee,
B.Pedrini,
and
K.Wüthrich
(2008).
NMR assignment of the nonstructural protein nsp3(1066-1181) from SARS-CoV.
|
| |
Biomol NMR Assign,
2,
135-138.
|
|
|
|
|
|
|
R.L.Graham,
J.S.Sparks,
L.D.Eckerle,
A.C.Sims,
and
M.R.Denison
(2008).
SARS coronavirus replicase proteins in pathogenesis.
|
| |
Virus Res,
133,
88.
|
|
|
|
|
|
|
Z.Li,
D.Barajas,
T.Panavas,
D.A.Herbst,
and
P.D.Nagy
(2008).
Cdc34p ubiquitin-conjugating enzyme is a component of the tombusvirus replicase complex and ubiquitinates p33 replication protein.
|
| |
J Virol,
82,
6911-6926.
|
|
|
|
|
|
|
A.Fernández-Montalván,
T.Bouwmeester,
G.Joberty,
R.Mader,
M.Mahnke,
B.Pierrat,
J.M.Schlaeppi,
S.Worpenberg,
and
B.Gerhartz
(2007).
Biochemical characterization of USP7 reveals post-translational modification sites and structural requirements for substrate processing and subcellular localization.
|
| |
FEBS J,
274,
4256-4270.
|
|
|
|
|
|
|
A.Kanjanahaluethai,
Z.Chen,
D.Jukneliene,
and
S.C.Baker
(2007).
Membrane topology of murine coronavirus replicase nonstructural protein 3.
|
| |
Virology,
361,
391-401.
|
|
|
|
|
|
|
C.Schlieker,
W.A.Weihofen,
E.Frijns,
L.M.Kattenhorn,
R.Gaudet,
and
H.L.Ploegh
(2007).
Structure of a herpesvirus-encoded cysteine protease reveals a unique class of deubiquitinating enzymes.
|
| |
Mol Cell,
25,
677-687.
|
|
|
PDB code:
|
|
|
|
|
|
|
|
J.Ziebuhr,
B.Schelle,
N.Karl,
E.Minskaia,
S.Bayer,
S.G.Siddell,
A.E.Gorbalenya,
and
V.Thiel
(2007).
Human coronavirus 229E papain-like proteases have overlapping specificities but distinct functions in viral replication.
|
| |
J Virol,
81,
3922-3932.
|
|
|
|
|
|
|
K.L.Maxwell,
and
L.Frappier
(2007).
Viral proteomics.
|
| |
Microbiol Mol Biol Rev,
71,
398-411.
|
|
|
|
|
|
|
M.Bartlam,
Y.Xu,
and
Z.Rao
(2007).
Structural proteomics of the SARS coronavirus: a model response to emerging infectious diseases.
|
| |
J Struct Funct Genomics,
8,
85-97.
|
|
|
|
|
|
|
M.S.Almeida,
M.A.Johnson,
T.Herrmann,
M.Geralt,
and
K.Wüthrich
(2007).
Novel beta-barrel fold in the nuclear magnetic resonance structure of the replicase nonstructural protein 1 from the severe acute respiratory syndrome coronavirus.
|
| |
J Virol,
81,
3151-3161.
|
|
|
PDB codes:
|
|
|
|
|
|
|
|
N.Frias-Staheli,
N.V.Giannakopoulos,
M.Kikkert,
S.L.Taylor,
A.Bridgen,
J.Paragas,
J.A.Richt,
R.R.Rowland,
C.S.Schmaljohn,
D.J.Lenschow,
E.J.Snijder,
A.García-Sastre,
and
H.W.Virgin
(2007).
Ovarian tumor domain-containing viral proteases evade ubiquitin- and ISG15-dependent innate immune responses.
|
| |
Cell Host Microbe,
2,
404-416.
|
|
|
|
|
|
|
P.Serrano,
M.A.Johnson,
M.S.Almeida,
R.Horst,
T.Herrmann,
J.S.Joseph,
B.W.Neuman,
V.Subramanian,
K.S.Saikatendu,
M.J.Buchmeier,
R.C.Stevens,
P.Kuhn,
and
K.Wüthrich
(2007).
Nuclear magnetic resonance structure of the N-terminal domain of nonstructural protein 3 from the severe acute respiratory syndrome coronavirus.
|
| |
J Virol,
81,
12049-12060.
|
|
|
PDB codes:
|
|
|
|
|
|
|
|
S.G.Devaraj,
N.Wang,
Z.Chen,
Z.Chen,
M.Tseng,
N.Barretto,
R.Lin,
C.J.Peters,
C.T.Tseng,
S.C.Baker,
and
K.Li
(2007).
Regulation of IRF-3-dependent innate immunity by the papain-like protease domain of the severe acute respiratory syndrome coronavirus.
|
| |
J Biol Chem,
282,
32208-32221.
|
|
|
|
|
|
|
S.G.Sawicki,
D.L.Sawicki,
and
S.G.Siddell
(2007).
A contemporary view of coronavirus transcription.
|
| |
J Virol,
81,
20-29.
|
|
|
|
|
|
|
V.C.Cheng,
S.K.Lau,
P.C.Woo,
and
K.Y.Yuen
(2007).
Severe acute respiratory syndrome coronavirus as an agent of emerging and reemerging infection.
|
| |
Clin Microbiol Rev,
20,
660-694.
|
|
|
|
|
|
|
X.Zhu,
R.Ménard,
and
T.Sulea
(2007).
High incidence of ubiquitin-like domains in human ubiquitin-specific proteases.
|
| |
Proteins,
69,
1-7.
|
|
|
|
|
|
|
Z.Chen,
Y.Wang,
K.Ratia,
A.D.Mesecar,
K.D.Wilkinson,
and
S.C.Baker
(2007).
Proteolytic processing and deubiquitinating activity of papain-like proteases of human coronavirus NL63.
|
| |
J Virol,
81,
6007-6018.
|
|
|
|
|
|
|
E.M.Hol,
D.F.Fischer,
H.Ovaa,
and
W.Scheper
(2006).
Ubiquitin proteasome system as a pharmacological target in neurodegeneration.
|
| |
Expert Rev Neurother,
6,
1337-1347.
|
|
|
|
|
|
|
H.Schütze,
R.Ulferts,
B.Schelle,
S.Bayer,
H.Granzow,
B.Hoffmann,
T.C.Mettenleiter,
and
J.Ziebuhr
(2006).
Characterization of White bream virus reveals a novel genetic cluster of nidoviruses.
|
| |
J Virol,
80,
11598-11609.
|
|
|
|
|
|
|
J.R.Mesters,
J.Tan,
and
R.Hilgenfeld
(2006).
Viral enzymes.
|
| |
Curr Opin Struct Biol,
16,
776-786.
|
|
|
|
|
|
|
M.Renatus,
S.G.Parrado,
A.D'Arcy,
U.Eidhoff,
B.Gerhartz,
U.Hassiepen,
B.Pierrat,
R.Riedl,
D.Vinzenz,
S.Worpenberg,
and
M.Kroemer
(2006).
Structural basis of ubiquitin recognition by the deubiquitinating protease USP2.
|
| |
Structure,
14,
1293-1302.
|
|
|
PDB code:
|
|
|
|
|
|
|
|
R.L.Graham,
and
M.R.Denison
(2006).
Replication of murine hepatitis virus is regulated by papain-like proteinase 1 processing of nonstructural proteins 1, 2, and 3.
|
| |
J Virol,
80,
11610-11620.
|
|
|
|
|
|
The most recent references are shown first.
Citation data come partly from CiteXplore and partly
from an automated harvesting procedure. Note that this is likely to be
only a partial list as not all journals are covered by
either method. However, we are continually building up the citation data
so more and more references will be included with time.
Where a reference describes a PDB structure, the PDB
code is
shown on the right.
|
|




 Links
Links








































