 |
PDBsum entry 2vj1

|
|
|
|
|
 |
Contents |
 |
|
|
|
|
|
|
|
|
|
|
|
|
* Residue conservation analysis
|
|
|
|
|
|
|
|
|
|
|
Enzyme class 2:
|
|
E.C.2.1.1.-
- ?????
|
|
|
|
|
|
|
Enzyme class 3:
|
|
E.C.2.1.1.56
- mRNA (guanine-N(7))-methyltransferase.
|
|
|
|
|
|
|
Reaction:
|
|
a 5'-end (5'-triphosphoguanosine)-ribonucleoside in mRNA + S-adenosyl-L- methionine = a 5'-end (N(7)-methyl 5'-triphosphoguanosine)-ribonucleoside in mRNA + S-adenosyl-L-homocysteine
|
|
|
|
|
|
5'-end (5'-triphosphoguanosine)-ribonucleoside in mRNA
|
+
|
 S-adenosyl-L- methionine
S-adenosyl-L- methionine
|
=
|
5'-end (N(7)-methyl 5'-triphosphoguanosine)-ribonucleoside in mRNA
|
+
|
 S-adenosyl-L-homocysteine
S-adenosyl-L-homocysteine
|
|
|
|
|
|
|
|
|
|
Enzyme class 4:
|
|
E.C.2.1.1.57
- methyltransferase cap1.
|
|
|
|
|
|
|
Reaction:
|
|
a 5'-end (N(7)-methyl 5'-triphosphoguanosine)-ribonucleoside in mRNA + S-adenosyl-L-methionine = a 5'-end (N(7)-methyl 5'-triphosphoguanosine)- (2'-O-methyl-ribonucleoside) in mRNA + S-adenosyl-L-homocysteine + H+
|
|
|
|
|
|
5'-end (N(7)-methyl 5'-triphosphoguanosine)-ribonucleoside in mRNA
|
+
|
S-adenosyl-L-methionine
|
=
|
5'-end (N(7)-methyl 5'-triphosphoguanosine)- (2'-O-methyl-ribonucleoside) in mRNA
|
+
|
S-adenosyl-L-homocysteine
|
+
|
H(+)
|
|
|
|
|
|
|
|
|
|
Enzyme class 5:
|
|
E.C.2.7.7.48
- RNA-directed Rna polymerase.
|
|
|
|
|
|
|
Reaction:
|
|
RNA(n) + a ribonucleoside 5'-triphosphate = RNA(n+1) + diphosphate
|
|
|
|
|
|
 RNA(n)
RNA(n)
|
+
|
ribonucleoside 5'-triphosphate
|
=
|
RNA(n+1)
|
+
|
 diphosphate
diphosphate
|
|
|
|
|
|
|
|
|
|
Enzyme class 6:
|
|
E.C.2.7.7.50
- mRNA guanylyltransferase.
|
|
|
|
|
|
|
Reaction:
|
|
a 5'-end diphospho-ribonucleoside in mRNA + GTP + H+ = a 5'-end (5'-triphosphoguanosine)-ribonucleoside in mRNA + diphosphate
|
|
|
|
|
|
5'-end diphospho-ribonucleoside in mRNA
|
+
|
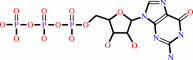 GTP
GTP
|
+
|
H(+)
|
=
|
5'-end (5'-triphosphoguanosine)-ribonucleoside in mRNA
|
+
|
diphosphate
|
|
|
|
|
|
|
|
|
|
Enzyme class 7:
|
|
E.C.3.1.13.-
- ?????
|
|
|
|
|
|
|
Enzyme class 8:
|
|
E.C.3.4.19.12
- ubiquitinyl hydrolase 1.
|
|
|
|
|
|
|
Reaction:
|
|
Thiol-dependent hydrolysis of ester, thiolester, amide, peptide and isopeptide bonds formed by the C-terminal Gly of ubiquitin (a 76-residue protein attached to proteins as an intracellular targeting signal).
|
|
|
|
|
|
Enzyme class 9:
|
|
E.C.3.4.22.-
- ?????
|
|
|
|
|
|
|
Enzyme class 10:
|
|
E.C.3.4.22.69
- Sars coronavirus main proteinase.
|
|
|
|
|
|
|
Enzyme class 11:
|
|
E.C.3.6.4.12
- Dna helicase.
|
|
|
|
|
|
|
Reaction:
|
|
ATP + H2O = ADP + phosphate + H+
|
|
|
|
|
|
 ATP
ATP
|
+
|
H2O
|
=
|
 ADP
ADP
|
+
|
 phosphate
phosphate
|
+
|
H(+)
|
|
|
|
|
|
|
|
|
|
Enzyme class 12:
|
|
E.C.3.6.4.13
- Rna helicase.
|
|
|
|
|
|
|
Reaction:
|
|
ATP + H2O = ADP + phosphate + H+
|
|
|
|
|
|
ATP
|
+
|
H2O
|
=
|
ADP
|
+
|
phosphate
|
+
|
H(+)
|
|
|
|
|
|
|
|
|
|
Enzyme class 13:
|
|
E.C.4.6.1.-
- ?????
|
|
|
|
|
|
|
|
|
|
Note, where more than one E.C. class is given (as above), each may
correspond to a different protein domain or, in the case of polyprotein
precursors, to a different mature protein.
|
|
|
|
Molecule diagrams generated from .mol files obtained from the
KEGG ftp site
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
| |
|
|
| |
|
|
Chem Biol
15:597-606
(2008)
|
|
PubMed id:
|
|
|
|
|
|
| |
|
A structural view of the inactivation of the SARS coronavirus main proteinase by benzotriazole esters.
|
|
K.H.Verschueren,
K.Pumpor,
S.Anemüller,
S.Chen,
J.R.Mesters,
R.Hilgenfeld.
|
|
|
|
|
| |
ABSTRACT
|
|
|
|
| |
|
|
The main proteinase (M(pro)) of the severe acute respiratory syndrome (SARS)
coronavirus is a principal target for the design of anticoronaviral compounds.
Benzotriazole esters have been reported as potent nonpeptidic inhibitors of the
enzyme, but their exact mechanism of action remains unclear. Here we present
crystal structures of SARS-CoV M(pro), the active-site cysteine of which has
been acylated by benzotriazole esters that act as suicide inhibitors. In one of
the structures, the thioester product has been hydrolyzed and benzoic acid is
observed to bind to the hydrophobic S2 pocket. This structure also features the
enzyme with a shortened N-terminal segment ("amputated N finger"). The results
further the understanding of the important role of the N finger for catalysis as
well as the design of benzotriazole inhibitors with improved specificity.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
 |
 |
|
Literature references that cite this PDB file's key reference
|
|
|
| |
PubMed id
|
|
Reference
|
|
|
|
|
|
A.K.Ghosh,
G.Gong,
V.Grum-Tokars,
D.C.Mulhearn,
S.C.Baker,
M.Coughlin,
B.S.Prabhakar,
K.Sleeman,
M.E.Johnson,
and
A.D.Mesecar
(2008).
Design, synthesis and antiviral efficacy of a series of potent chloropyridyl ester-derived SARS-CoV 3CLpro inhibitors.
|
| |
Bioorg Med Chem Lett,
18,
5684-5688.
|
|
|
|
|
|
The most recent references are shown first.
Citation data come partly from CiteXplore and partly
from an automated harvesting procedure. Note that this is likely to be
only a partial list as not all journals are covered by
either method. However, we are continually building up the citation data
so more and more references will be included with time.
|
|


 Links
Links






































