 |
PDBsum entry 2qcy

|
|
|
|
|
 |
Contents |
 |
|
|
|
|
|
|
|
|
|
|
* Residue conservation analysis
|
|
|
|
|
|
|
|
|
|
|
Enzyme class 2:
|
|
E.C.2.1.1.-
- ?????
|
|
|
|
|
|
|
Enzyme class 3:
|
|
E.C.2.1.1.56
- mRNA (guanine-N(7))-methyltransferase.
|
|
|
|
|
|
|
Reaction:
|
|
a 5'-end (5'-triphosphoguanosine)-ribonucleoside in mRNA + S-adenosyl-L- methionine = a 5'-end (N(7)-methyl 5'-triphosphoguanosine)-ribonucleoside in mRNA + S-adenosyl-L-homocysteine
|
|
|
|
|
|
5'-end (5'-triphosphoguanosine)-ribonucleoside in mRNA
|
+
|
 S-adenosyl-L- methionine
S-adenosyl-L- methionine
|
=
|
5'-end (N(7)-methyl 5'-triphosphoguanosine)-ribonucleoside in mRNA
|
+
|
 S-adenosyl-L-homocysteine
S-adenosyl-L-homocysteine
|
|
|
|
|
|
|
|
|
|
Enzyme class 4:
|
|
E.C.2.1.1.57
- methyltransferase cap1.
|
|
|
|
|
|
|
Reaction:
|
|
a 5'-end (N(7)-methyl 5'-triphosphoguanosine)-ribonucleoside in mRNA + S-adenosyl-L-methionine = a 5'-end (N(7)-methyl 5'-triphosphoguanosine)- (2'-O-methyl-ribonucleoside) in mRNA + S-adenosyl-L-homocysteine + H+
|
|
|
|
|
|
5'-end (N(7)-methyl 5'-triphosphoguanosine)-ribonucleoside in mRNA
|
+
|
S-adenosyl-L-methionine
|
=
|
5'-end (N(7)-methyl 5'-triphosphoguanosine)- (2'-O-methyl-ribonucleoside) in mRNA
|
+
|
S-adenosyl-L-homocysteine
|
+
|
H(+)
|
|
|
|
|
|
|
|
|
|
Enzyme class 5:
|
|
E.C.2.7.7.48
- RNA-directed Rna polymerase.
|
|
|
|
|
|
|
Reaction:
|
|
RNA(n) + a ribonucleoside 5'-triphosphate = RNA(n+1) + diphosphate
|
|
|
|
|
|
 RNA(n)
RNA(n)
|
+
|
ribonucleoside 5'-triphosphate
|
=
|
RNA(n+1)
|
+
|
 diphosphate
diphosphate
|
|
|
|
|
|
|
|
|
|
Enzyme class 6:
|
|
E.C.2.7.7.50
- mRNA guanylyltransferase.
|
|
|
|
|
|
|
Reaction:
|
|
a 5'-end diphospho-ribonucleoside in mRNA + GTP + H+ = a 5'-end (5'-triphosphoguanosine)-ribonucleoside in mRNA + diphosphate
|
|
|
|
|
|
5'-end diphospho-ribonucleoside in mRNA
|
+
|
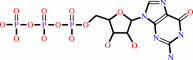 GTP
GTP
|
+
|
H(+)
|
=
|
5'-end (5'-triphosphoguanosine)-ribonucleoside in mRNA
|
+
|
diphosphate
|
|
|
|
|
|
|
|
|
|
Enzyme class 7:
|
|
E.C.3.1.13.-
- ?????
|
|
|
|
|
|
|
Enzyme class 8:
|
|
E.C.3.4.19.12
- ubiquitinyl hydrolase 1.
|
|
|
|
|
|
|
Reaction:
|
|
Thiol-dependent hydrolysis of ester, thiolester, amide, peptide and isopeptide bonds formed by the C-terminal Gly of ubiquitin (a 76-residue protein attached to proteins as an intracellular targeting signal).
|
|
|
|
|
|
Enzyme class 9:
|
|
E.C.3.4.22.-
- ?????
|
|
|
|
|
|
|
Enzyme class 10:
|
|
E.C.3.4.22.69
- Sars coronavirus main proteinase.
|
|
|
|
|
|
|
Enzyme class 11:
|
|
E.C.3.6.4.12
- Dna helicase.
|
|
|
|
|
|
|
Reaction:
|
|
ATP + H2O = ADP + phosphate + H+
|
|
|
|
|
|
 ATP
ATP
|
+
|
H2O
|
=
|
 ADP
ADP
|
+
|
 phosphate
phosphate
|
+
|
H(+)
|
|
|
|
|
|
|
|
|
|
Enzyme class 12:
|
|
E.C.3.6.4.13
- Rna helicase.
|
|
|
|
|
|
|
Reaction:
|
|
ATP + H2O = ADP + phosphate + H+
|
|
|
|
|
|
ATP
|
+
|
H2O
|
=
|
ADP
|
+
|
phosphate
|
+
|
H(+)
|
|
|
|
|
|
|
|
|
|
Enzyme class 13:
|
|
E.C.4.6.1.-
- ?????
|
|
|
|
|
|
|
|
|
|
Note, where more than one E.C. class is given (as above), each may
correspond to a different protein domain or, in the case of polyprotein
precursors, to a different mature protein.
|
|
|
|
Molecule diagrams generated from .mol files obtained from the
KEGG ftp site
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
| |
|
|
| |
|
|
J Virol
82:4620-4629
(2008)
|
|
PubMed id:
|
|
|
|
|
|
| |
|
Mechanism for controlling the dimer-monomer switch and coupling dimerization to catalysis of the severe acute respiratory syndrome coronavirus 3C-like protease.
|
|
J.Shi,
J.Sivaraman,
J.Song.
|
|
|
|
|
| |
ABSTRACT
|
|
|
|
| |
|
|
Unlike 3C protease, the severe acute respiratory syndrome coronavirus (SARS-CoV)
3C-like protease (3CLpro) is only enzymatically active as a homodimer and its
catalysis is under extensive regulation by the unique extra domain. Despite
intense studies, two puzzles still remain: (i) how the dimer-monomer switch is
controlled and (ii) why dimerization is absolutely required for catalysis. Here
we report the monomeric crystal structure of the SARS-CoV 3CLpro mutant R298A at
a resolution of 1.75 A. Detailed analysis reveals that Arg298 serves as a key
component for maintaining dimerization, and consequently, its mutation will
trigger a cooperative switch from a dimer to a monomer. The monomeric enzyme is
irreversibly inactivated because its catalytic machinery is frozen in the
collapsed state, characteristic of the formation of a short 3(10)-helix from an
active-site loop. Remarkably, dimerization appears to be coupled to catalysis in
3CLpro through the use of overlapped residues for two networks, one for
dimerization and another for the catalysis.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
 |
 |
|
Literature references that cite this PDB file's key reference
|
|
|
| |
PubMed id
|
|
Reference
|
|
|
|
|
|
J.Shi,
N.Han,
L.Lim,
S.Lua,
J.Sivaraman,
L.Wang,
Y.Mu,
and
J.Song
(2011).
Dynamically-driven inactivation of the catalytic machinery of the SARS 3C-like protease by the N214A mutation on the extra domain.
|
| |
PLoS Comput Biol,
7,
e1001084.
|
|
|
|
|
|
|
M.Y.Tsai,
W.H.Chang,
J.Y.Liang,
L.L.Lin,
G.G.Chang,
and
H.P.Chang
(2010).
Essential covalent linkage between the chymotrypsin-like domain and the extra domain of the SARS-CoV main protease.
|
| |
J Biochem,
148,
349-358.
|
|
|
|
|
|
|
S.C.Cheng,
G.G.Chang,
and
C.Y.Chou
(2010).
Mutation of Glu-166 blocks the substrate-induced dimerization of SARS coronavirus main protease.
|
| |
Biophys J,
98,
1327-1336.
|
|
|
|
|
|
|
H.Qin,
J.Shi,
R.Noberini,
E.B.Pasquale,
and
J.Song
(2008).
Crystal structure and NMR binding reveal that two small molecule antagonists target the high affinity ephrin-binding channel of the EphA4 receptor.
|
| |
J Biol Chem,
283,
29473-29484.
|
|
|
PDB code:
|
|
|
|
|
|
|
The most recent references are shown first.
Citation data come partly from CiteXplore and partly
from an automated harvesting procedure. Note that this is likely to be
only a partial list as not all journals are covered by
either method. However, we are continually building up the citation data
so more and more references will be included with time.
Where a reference describes a PDB structure, the PDB
code is
shown on the right.
|
|


 Links
Links






































