 |
PDBsum entry 2op9

|
|
|
|
|
 |
Contents |
 |
|
|
|
|
|
|
|
|
|
|
|
|
* Residue conservation analysis
|
|
|
|
|
|
|
|
|
|
|
Enzyme class 2:
|
|
E.C.2.1.1.-
- ?????
|
|
|
|
|
|
|
Enzyme class 3:
|
|
E.C.2.1.1.56
- mRNA (guanine-N(7))-methyltransferase.
|
|
|
|
|
|
|
Reaction:
|
|
a 5'-end (5'-triphosphoguanosine)-ribonucleoside in mRNA + S-adenosyl-L- methionine = a 5'-end (N(7)-methyl 5'-triphosphoguanosine)-ribonucleoside in mRNA + S-adenosyl-L-homocysteine
|
|
|
|
|
|
5'-end (5'-triphosphoguanosine)-ribonucleoside in mRNA
|
+
|
 S-adenosyl-L- methionine
S-adenosyl-L- methionine
|
=
|
5'-end (N(7)-methyl 5'-triphosphoguanosine)-ribonucleoside in mRNA
|
+
|
 S-adenosyl-L-homocysteine
S-adenosyl-L-homocysteine
|
|
|
|
|
|
|
|
|
|
Enzyme class 4:
|
|
E.C.2.1.1.57
- methyltransferase cap1.
|
|
|
|
|
|
|
Reaction:
|
|
a 5'-end (N(7)-methyl 5'-triphosphoguanosine)-ribonucleoside in mRNA + S-adenosyl-L-methionine = a 5'-end (N(7)-methyl 5'-triphosphoguanosine)- (2'-O-methyl-ribonucleoside) in mRNA + S-adenosyl-L-homocysteine + H+
|
|
|
|
|
|
5'-end (N(7)-methyl 5'-triphosphoguanosine)-ribonucleoside in mRNA
|
+
|
S-adenosyl-L-methionine
|
=
|
5'-end (N(7)-methyl 5'-triphosphoguanosine)- (2'-O-methyl-ribonucleoside) in mRNA
|
+
|
S-adenosyl-L-homocysteine
|
+
|
H(+)
|
|
|
|
|
|
|
|
|
|
Enzyme class 5:
|
|
E.C.2.7.7.48
- RNA-directed Rna polymerase.
|
|
|
|
|
|
|
Reaction:
|
|
RNA(n) + a ribonucleoside 5'-triphosphate = RNA(n+1) + diphosphate
|
|
|
|
|
|
 RNA(n)
RNA(n)
|
+
|
ribonucleoside 5'-triphosphate
|
=
|
RNA(n+1)
|
+
|
 diphosphate
diphosphate
|
|
|
|
|
|
|
|
|
|
Enzyme class 6:
|
|
E.C.2.7.7.50
- mRNA guanylyltransferase.
|
|
|
|
|
|
|
Reaction:
|
|
a 5'-end diphospho-ribonucleoside in mRNA + GTP + H+ = a 5'-end (5'-triphosphoguanosine)-ribonucleoside in mRNA + diphosphate
|
|
|
|
|
|
5'-end diphospho-ribonucleoside in mRNA
|
+
|
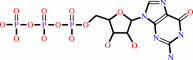 GTP
GTP
|
+
|
H(+)
|
=
|
5'-end (5'-triphosphoguanosine)-ribonucleoside in mRNA
|
+
|
diphosphate
|
|
|
|
|
|
|
|
|
|
Enzyme class 7:
|
|
E.C.3.1.13.-
- ?????
|
|
|
|
|
|
|
Enzyme class 8:
|
|
E.C.3.4.19.12
- ubiquitinyl hydrolase 1.
|
|
|
|
|
|
|
Reaction:
|
|
Thiol-dependent hydrolysis of ester, thiolester, amide, peptide and isopeptide bonds formed by the C-terminal Gly of ubiquitin (a 76-residue protein attached to proteins as an intracellular targeting signal).
|
|
|
|
|
|
Enzyme class 9:
|
|
E.C.3.4.22.-
- ?????
|
|
|
|
|
|
|
Enzyme class 10:
|
|
E.C.3.4.22.69
- Sars coronavirus main proteinase.
|
|
|
|
|
|
|
Enzyme class 11:
|
|
E.C.3.6.4.12
- Dna helicase.
|
|
|
|
|
|
|
Reaction:
|
|
ATP + H2O = ADP + phosphate + H+
|
|
|
|
|
|
 ATP
ATP
|
+
|
H2O
|
=
|
 ADP
ADP
|
+
|
 phosphate
phosphate
|
+
|
H(+)
|
|
|
|
|
|
|
|
|
|
Enzyme class 12:
|
|
E.C.3.6.4.13
- Rna helicase.
|
|
|
|
|
|
|
Reaction:
|
|
ATP + H2O = ADP + phosphate + H+
|
|
|
|
|
|
ATP
|
+
|
H2O
|
=
|
ADP
|
+
|
phosphate
|
+
|
H(+)
|
|
|
|
|
|
|
|
|
|
Enzyme class 13:
|
|
E.C.4.6.1.-
- ?????
|
|
|
|
|
|
|
|
|
|
Note, where more than one E.C. class is given (as above), each may
correspond to a different protein domain or, in the case of polyprotein
precursors, to a different mature protein.
|
|
|
|
Molecule diagrams generated from .mol files obtained from the
KEGG ftp site
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
| |
|
|
| |
|
|
Biochemistry
46:8744-8752
(2007)
|
|
PubMed id:
|
|
|
|
|
|
| |
|
Substrate specificity profiling and identification of a new class of inhibitor for the major protease of the SARS coronavirus.
|
|
D.H.Goetz,
Y.Choe,
E.Hansell,
Y.T.Chen,
M.McDowell,
C.B.Jonsson,
W.R.Roush,
J.McKerrow,
C.S.Craik.
|
|
|
|
|
| |
ABSTRACT
|
|
|
|
| |
|
|
Severe acute respiratory syndrome (SARS) is an emerging infectious disease
associated with a high rate of mortality. The SARS-associated coronavirus
(SARS-CoV) has been identified as the etiological agent of the disease. Although
public health procedures have been effective in combating the spread of SARS,
concern remains about the possibility of a recurrence. Various approaches are
being pursued for the development of efficacious therapeutics. One promising
approach is to develop small molecule inhibitors of the essential major
polyprotein processing protease 3Clpro. Here we report a complete description of
the tetrapeptide substrate specificity of 3Clpro using fully degenerate peptide
libraries consisting of all 160,000 possible naturally occurring tetrapeptides.
The substrate specificity data show the expected P1-Gln P2-Leu specificity and
elucidate a novel preference for P1-His containing substrates equal to the
expected preference for P1-Gln. These data were then used to develop optimal
substrates for a high-throughput screen of a 2000 compound small-molecule
inhibitor library consisting of known cysteine protease inhibitor scaffolds. We
also report the 1.8 A X-ray crystal structure of 3Clpro bound to an irreversible
inhibitor. This inhibitor, an alpha,beta-epoxyketone, inhibits 3Clpro with a
k3/Ki of 0.002 microM(-1) s(-1) in a mode consistent with the substrate
specificity data. Finally, we report the successful rational improvement of this
scaffold with second generation inhibitors. These data provide the foundation
for a rational small-molecule inhibitor design effort based upon the inhibitor
scaffold identified, the crystal structure of the complex, and a more complete
understanding of P1-P4 substrate specificity.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
 |
 |
|
Literature references that cite this PDB file's key reference
|
|
|
| |
PubMed id
|
|
Reference
|
|
|
|
|
|
C.P.Chuck,
L.T.Chong,
C.Chen,
H.F.Chow,
D.C.Wan,
and
K.B.Wong
(2010).
Profiling of substrate specificity of SARS-CoV 3CL.
|
| |
PLoS One,
5,
e13197.
|
|
|
|
|
|
|
D.N.Okamoto,
L.C.Oliveira,
M.Y.Kondo,
M.H.Cezari,
Z.Szeltner,
T.Juhász,
M.A.Juliano,
L.Polgár,
L.Juliano,
and
I.E.Gouvea
(2010).
Increase of SARS-CoV 3CL peptidase activity due to macromolecular crowding effects in the milieu composition.
|
| |
Biol Chem,
391,
1461-1468.
|
|
|
|
|
|
|
S.Fang,
H.Shen,
J.Wang,
F.P.Tay,
and
D.X.Liu
(2010).
Functional and genetic studies of the substrate specificity of coronavirus infectious bronchitis virus 3C-like proteinase.
|
| |
J Virol,
84,
7325-7336.
|
|
|
|
|
|
|
M.D.Lim,
and
C.S.Craik
(2009).
Using specificity to strategically target proteases.
|
| |
Bioorg Med Chem,
17,
1094-1100.
|
|
|
|
|
|
|
S.Subramanian,
M.Hardt,
Y.Choe,
R.K.Niles,
E.B.Johansen,
J.Legac,
J.Gut,
I.D.Kerr,
C.S.Craik,
and
P.J.Rosenthal
(2009).
Hemoglobin cleavage site-specificity of the Plasmodium falciparum cysteine proteases falcipain-2 and falcipain-3.
|
| |
PLoS ONE,
4,
e5156.
|
|
|
|
|
|
|
J.S.Sparks,
E.F.Donaldson,
X.Lu,
R.S.Baric,
and
M.R.Denison
(2008).
A novel mutation in murine hepatitis virus nsp5, the viral 3C-like proteinase, causes temperature-sensitive defects in viral growth and protein processing.
|
| |
J Virol,
82,
5999-6008.
|
|
|
|
|
|
|
N.Green,
R.D.Ott,
R.J.Isaacs,
and
H.Fang
(2008).
Cell-based Assays to Identify Inhibitors of Viral Disease.
|
| |
Expert Opin Drug Discov,
3,
671-676.
|
|
|
|
|
|
The most recent references are shown first.
Citation data come partly from CiteXplore and partly
from an automated harvesting procedure. Note that this is likely to be
only a partial list as not all journals are covered by
either method. However, we are continually building up the citation data
so more and more references will be included with time.
|
|


 Links
Links






































