 |
PDBsum entry 2fav

|
|
|
|
|
|

|

|

|

|
|

|
|

|
|

|
|

|
|

|
|
|
 |

|

|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Viral protein
|
PDB id
|
|
|
|
2fav
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|

|

|

|
|
|
|
|
|
|
|
 |
Contents |
 |
|
|
|
|
|
|
|
|
|
|
|
|
* Residue conservation analysis
|
|
|
|
|
|
|
|
|
|
|
Enzyme class 2:
|
|
E.C.2.1.1.-
- ?????
|
|
|
|
|
|
|
Enzyme class 3:
|
|
E.C.2.1.1.56
- mRNA (guanine-N(7))-methyltransferase.
|
|
|
|
|
|
|
Reaction:
|
|
a 5'-end (5'-triphosphoguanosine)-ribonucleoside in mRNA + S-adenosyl-L- methionine = a 5'-end (N(7)-methyl 5'-triphosphoguanosine)-ribonucleoside in mRNA + S-adenosyl-L-homocysteine
|
|
|
|
|
|
5'-end (5'-triphosphoguanosine)-ribonucleoside in mRNA
|
+
|
 S-adenosyl-L- methionine
S-adenosyl-L- methionine
|
=
|
5'-end (N(7)-methyl 5'-triphosphoguanosine)-ribonucleoside in mRNA
|
+
|
 S-adenosyl-L-homocysteine
S-adenosyl-L-homocysteine
|
|
|
|
|
|
|
|
|
|
Enzyme class 4:
|
|
E.C.2.1.1.57
- methyltransferase cap1.
|
|
|
|
|
|
|
Reaction:
|
|
a 5'-end (N(7)-methyl 5'-triphosphoguanosine)-ribonucleoside in mRNA + S-adenosyl-L-methionine = a 5'-end (N(7)-methyl 5'-triphosphoguanosine)- (2'-O-methyl-ribonucleoside) in mRNA + S-adenosyl-L-homocysteine + H+
|
|
|
|
|
|
5'-end (N(7)-methyl 5'-triphosphoguanosine)-ribonucleoside in mRNA
|
+
|
S-adenosyl-L-methionine
|
=
|
5'-end (N(7)-methyl 5'-triphosphoguanosine)- (2'-O-methyl-ribonucleoside) in mRNA
|
+
|
S-adenosyl-L-homocysteine
|
+
|
H(+)
|
|
|
|
|
|
|
|
|
|
Enzyme class 5:
|
|
E.C.2.7.7.48
- RNA-directed Rna polymerase.
|
|
|
|
|
|
|
Reaction:
|
|
RNA(n) + a ribonucleoside 5'-triphosphate = RNA(n+1) + diphosphate
|
|
|
|
|
|
 RNA(n)
RNA(n)
|
+
|
ribonucleoside 5'-triphosphate
|
=
|
RNA(n+1)
Bound ligand (Het Group name = )
matches with 75.00% similarity
|
+
|
 diphosphate
diphosphate
|
|
|
|
|
|
|
|
|
|
Enzyme class 6:
|
|
E.C.2.7.7.50
- mRNA guanylyltransferase.
|
|
|
|
|
|
|
Reaction:
|
|
a 5'-end diphospho-ribonucleoside in mRNA + GTP + H+ = a 5'-end (5'-triphosphoguanosine)-ribonucleoside in mRNA + diphosphate
|
|
|
|
|
|
5'-end diphospho-ribonucleoside in mRNA
|
+
|
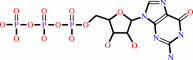 GTP
GTP
|
+
|
H(+)
|
=
|
5'-end (5'-triphosphoguanosine)-ribonucleoside in mRNA
|
+
|
diphosphate
|
|
|
|
|
|
|
|
|
|
Enzyme class 7:
|
|
E.C.3.1.13.-
- ?????
|
|
|
|
|
|
|
Enzyme class 8:
|
|
E.C.3.4.19.12
- ubiquitinyl hydrolase 1.
|
|
|
|
|
|
|
Reaction:
|
|
Thiol-dependent hydrolysis of ester, thiolester, amide, peptide and isopeptide bonds formed by the C-terminal Gly of ubiquitin (a 76-residue protein attached to proteins as an intracellular targeting signal).
|
|
|
|
|
|
Enzyme class 9:
|
|
E.C.3.4.22.-
- ?????
|
|
|
|
|
|
|
Enzyme class 10:
|
|
E.C.3.4.22.69
- Sars coronavirus main proteinase.
|
|
|
|
|
|
|
Enzyme class 11:
|
|
E.C.3.6.4.12
- Dna helicase.
|
|
|
|
|
|
|
Reaction:
|
|
ATP + H2O = ADP + phosphate + H+
|
|
|
|
|
|
 ATP
ATP
|
+
|
H2O
|
=
|
 ADP
ADP
|
+
|
 phosphate
phosphate
|
+
|
H(+)
|
|
|
|
|
|
|
|
|
|
Enzyme class 12:
|
|
E.C.3.6.4.13
- Rna helicase.
|
|
|
|
|
|
|
Reaction:
|
|
ATP + H2O = ADP + phosphate + H+
|
|
|
|
|
|
ATP
|
+
|
H2O
|
=
|
ADP
|
+
|
phosphate
|
+
|
H(+)
|
|
|
|
|
|
|
|
|
|
Enzyme class 13:
|
|
E.C.4.6.1.-
- ?????
|
|
|
|
|
|
|
|
|
|
Note, where more than one E.C. class is given (as above), each may
correspond to a different protein domain or, in the case of polyprotein
precursors, to a different mature protein.
|
|
|
|
Molecule diagrams generated from .mol files obtained from the
KEGG ftp site
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
| |
|
|
| |
|
|
J Virol
80:8493-8502
(2006)
|
|
PubMed id:
|
|
|
|
|
|
| |
|
Structural and functional basis for ADP-ribose and poly(ADP-ribose) binding by viral macro domains.
|
|
M.P.Egloff,
H.Malet,
A.Putics,
M.Heinonen,
H.Dutartre,
A.Frangeul,
A.Gruez,
V.Campanacci,
C.Cambillau,
J.Ziebuhr,
T.Ahola,
B.Canard.
|
|
|
|
|
| |
ABSTRACT
|
|
|
|
| |
|
|
Macro domains constitute a protein module family found associated with specific
histones and proteins involved in chromatin metabolism. In addition, a small
number of animal RNA viruses, such as corona- and toroviruses, alphaviruses, and
hepatitis E virus, encode macro domains for which, however, structural and
functional information is extremely limited. Here, we characterized the macro
domains from hepatitis E virus, Semliki Forest virus, and severe acute
respiratory syndrome coronavirus (SARS-CoV). The crystal structure of the
SARS-CoV macro domain was determined at 1.8-Angstroms resolution in complex with
ADP-ribose. Information derived from structural, mutational, and sequence
analyses suggests a close phylogenetic and, most probably, functional
relationship between viral and cellular macro domain homologs. The data revealed
that viral macro domains have relatively poor ADP-ribose 1"-phosphohydrolase
activities (which were previously proposed to be their biologically relevant
function) but bind efficiently free and poly(ADP-ribose) polymerase 1-bound
poly(ADP-ribose) in vitro. Collectively, these results suggest to further
evaluate the role of viral macro domains in host response to viral infection.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
 |
 |
|
Literature references that cite this PDB file's key reference
|
|
|
| |
PubMed id
|
|
Reference
|
|
|
|
|
|
B.F.Beitzel,
R.R.Bakken,
J.M.Smith,
and
C.S.Schmaljohn
(2010).
High-resolution functional mapping of the venezuelan equine encephalitis virus genome by insertional mutagenesis and massively parallel sequencing.
|
| |
PLoS Pathog,
6,
e1001146.
|
|
|
|
|
|
|
H.L.Stokes,
S.Baliji,
C.G.Hui,
S.G.Sawicki,
S.C.Baker,
and
S.G.Siddell
(2010).
A new cistron in the murine hepatitis virus replicase gene.
|
| |
J Virol,
84,
10148-10158.
|
|
|
|
|
|
|
K.R.Hurst,
R.Ye,
S.J.Goebel,
P.Jayaraman,
and
P.S.Masters
(2010).
An interaction between the nucleocapsid protein and a component of the replicase-transcriptase complex is crucial for the infectivity of coronavirus genomic RNA.
|
| |
J Virol,
84,
10276-10288.
|
|
|
|
|
|
|
M.Varjak,
E.Zusinaite,
and
A.Merits
(2010).
Novel functions of the alphavirus nonstructural protein nsP3 C-terminal region.
|
| |
J Virol,
84,
2352-2364.
|
|
|
|
|
|
|
S.J.Bender,
and
S.R.Weiss
(2010).
Pathogenesis of murine coronavirus in the central nervous system.
|
| |
J Neuroimmune Pharmacol,
5,
336-354.
|
|
|
|
|
|
|
Y.A.Karpe,
and
K.S.Lole
(2010).
NTPase and 5' to 3' RNA duplex-unwinding activities of the hepatitis E virus helicase domain.
|
| |
J Virol,
84,
3595-3602.
|
|
|
|
|
|
|
A.Chatterjee,
M.A.Johnson,
P.Serrano,
B.Pedrini,
J.S.Joseph,
B.W.Neuman,
K.Saikatendu,
M.J.Buchmeier,
P.Kuhn,
and
K.Wüthrich
(2009).
Nuclear magnetic resonance structure shows that the severe acute respiratory syndrome coronavirus-unique domain contains a macrodomain fold.
|
| |
J Virol,
83,
1823-1836.
|
|
|
PDB codes:
|
|
|
|
|
|
|
|
E.Park,
and
D.E.Griffin
(2009).
Interaction of Sindbis virus non-structural protein 3 with poly(ADP-ribose) polymerase 1 in neuronal cells.
|
| |
J Gen Virol,
90,
2073-2080.
|
|
|
|
|
|
|
E.Park,
and
D.E.Griffin
(2009).
The nsP3 macro domain is important for Sindbis virus replication in neurons and neurovirulence in mice.
|
| |
Virology,
388,
305-314.
|
|
|
|
|
|
|
H.A.Basta,
S.B.Cleveland,
R.A.Clinton,
A.G.Dimitrov,
and
M.A.McClure
(2009).
Evolution of teleost fish retroviruses: characterization of new retroviruses with cellular genes.
|
| |
J Virol,
83,
10152-10162.
|
|
|
|
|
|
|
H.Malet,
B.Coutard,
S.Jamal,
H.Dutartre,
N.Papageorgiou,
M.Neuvonen,
T.Ahola,
N.Forrester,
E.A.Gould,
D.Lafitte,
F.Ferron,
J.Lescar,
A.E.Gorbalenya,
X.de Lamballerie,
and
B.Canard
(2009).
The crystal structures of Chikungunya and Venezuelan equine encephalitis virus nsP3 macro domains define a conserved adenosine binding pocket.
|
| |
J Virol,
83,
6534-6545.
|
|
|
PDB codes:
|
|
|
|
|
|
|
|
J.A.Wojdyla,
I.Manolaridis,
E.J.Snijder,
A.E.Gorbalenya,
B.Coutard,
Y.Piotrowski,
R.Hilgenfeld,
and
P.A.Tucker
(2009).
Structure of the X (ADRP) domain of nsp3 from feline coronavirus.
|
| |
Acta Crystallogr D Biol Crystallogr,
65,
1292-1300.
|
|
|
PDB codes:
|
|
|
|
|
|
|
|
J.Tan,
C.Vonrhein,
O.S.Smart,
G.Bricogne,
M.Bollati,
Y.Kusov,
G.Hansen,
J.R.Mesters,
C.L.Schmidt,
and
R.Hilgenfeld
(2009).
The SARS-Unique Domain (SUD) of SARS Coronavirus Contains Two Macrodomains That Bind G-Quadruplexes.
|
| |
PLoS Pathog,
5,
e1000428.
|
|
|
PDB codes:
|
|
|
|
|
|
|
|
K.M.Rose,
and
S.R.Weiss
(2009).
Murine Coronavirus Cell Type Dependent Interaction with the Type I Interferon Response.
|
| |
Viruses,
1,
689-712.
|
|
|
|
|
|
|
P.Serrano,
M.A.Johnson,
A.Chatterjee,
B.W.Neuman,
J.S.Joseph,
M.J.Buchmeier,
P.Kuhn,
and
K.Wüthrich
(2009).
Nuclear magnetic resonance structure of the nucleic acid-binding domain of severe acute respiratory syndrome coronavirus nonstructural protein 3.
|
| |
J Virol,
83,
12998-13008.
|
|
|
PDB code:
|
|
|
|
|
|
|
|
Y.Piotrowski,
G.Hansen,
A.L.Boomaars-van der Zanden,
E.J.Snijder,
A.E.Gorbalenya,
and
R.Hilgenfeld
(2009).
Crystal structures of the X-domains of a Group-1 and a Group-3 coronavirus reveal that ADP-ribose-binding may not be a conserved property.
|
| |
Protein Sci,
18,
6.
|
|
|
PDB codes:
|
|
|
|
|
|
|
|
Y.Xu,
L.Cong,
C.Chen,
L.Wei,
Q.Zhao,
X.Xu,
Y.Ma,
M.Bartlam,
and
Z.Rao
(2009).
Crystal structures of two coronavirus ADP-ribose-1''-monophosphatases and their complexes with ADP-Ribose: a systematic structural analysis of the viral ADRP domain.
|
| |
J Virol,
83,
1083-1092.
|
|
|
PDB codes:
|
|
|
|
|
|
|
|
A.Hakmé,
A.Huber,
P.Dollé,
and
V.Schreiber
(2008).
The macroPARP genes parp-9 and parp-14 are developmentally and differentially regulated in mouse tissues.
|
| |
Dev Dyn,
237,
209-215.
|
|
|
|
|
|
|
A.Hakmé,
H.K.Wong,
F.Dantzer,
and
V.Schreiber
(2008).
The expanding field of poly(ADP-ribosyl)ation reactions. 'Protein Modifications: Beyond the Usual Suspects' Review Series.
|
| |
EMBO Rep,
9,
1094-1100.
|
|
|
|
|
|
|
J.P.Gagné,
M.Isabelle,
K.S.Lo,
S.Bourassa,
M.J.Hendzel,
V.L.Dawson,
T.M.Dawson,
and
G.G.Poirier
(2008).
Proteome-wide identification of poly(ADP-ribose) binding proteins and poly(ADP-ribose)-associated protein complexes.
|
| |
Nucleic Acids Res,
36,
6959-6976.
|
|
|
|
|
|
|
K.K.Eriksson,
L.Cervantes-Barragán,
B.Ludewig,
and
V.Thiel
(2008).
Mouse hepatitis virus liver pathology is dependent on ADP-ribose-1''-phosphatase, a viral function conserved in the alpha-like supergroup.
|
| |
J Virol,
82,
12325-12334.
|
|
|
|
|
|
|
M.Bartlam,
X.Xue,
and
Z.Rao
(2008).
The search for a structural basis for therapeutic intervention against the SARS coronavirus.
|
| |
Acta Crystallogr A,
64,
204-213.
|
|
|
|
|
|
|
O.Okhrimenko,
and
I.Jelesarov
(2008).
A survey of the year 2006 literature on applications of isothermal titration calorimetry.
|
| |
J Mol Recognit,
21,
1.
|
|
|
|
|
|
|
V.Lulla,
D.L.Sawicki,
S.G.Sawicki,
A.Lulla,
A.Merits,
and
T.Ahola
(2008).
Molecular defects caused by temperature-sensitive mutations in Semliki Forest virus nsP1.
|
| |
J Virol,
82,
9236-9244.
|
|
|
|
|
|
|
J.Ziebuhr,
B.Schelle,
N.Karl,
E.Minskaia,
S.Bayer,
S.G.Siddell,
A.E.Gorbalenya,
and
V.Thiel
(2007).
Human coronavirus 229E papain-like proteases have overlapping specificities but distinct functions in viral replication.
|
| |
J Virol,
81,
3922-3932.
|
|
|
|
|
|
|
M.Bartlam,
Y.Xu,
and
Z.Rao
(2007).
Structural proteomics of the SARS coronavirus: a model response to emerging infectious diseases.
|
| |
J Struct Funct Genomics,
8,
85-97.
|
|
|
|
|
|
|
S.Mattiussi,
I.Tempera,
G.Matusali,
G.Mearini,
L.Lenti,
S.Fratarcangeli,
L.Mosca,
M.D'Erme,
and
E.Mattia
(2007).
Inhibition of Poly(ADP-ribose)polymerase impairs Epstein Barr Virus lytic cycle progression.
|
| |
Infect Agent Cancer,
2,
18.
|
|
|
|
|
|
|
H.Schütze,
R.Ulferts,
B.Schelle,
S.Bayer,
H.Granzow,
B.Hoffmann,
T.C.Mettenleiter,
and
J.Ziebuhr
(2006).
Characterization of White bream virus reveals a novel genetic cluster of nidoviruses.
|
| |
J Virol,
80,
11598-11609.
|
|
|
|
|
|
|
J.R.Mesters,
J.Tan,
and
R.Hilgenfeld
(2006).
Viral enzymes.
|
| |
Curr Opin Struct Biol,
16,
776-786.
|
|
|
|
|
|
The most recent references are shown first.
Citation data come partly from CiteXplore and partly
from an automated harvesting procedure. Note that this is likely to be
only a partial list as not all journals are covered by
either method. However, we are continually building up the citation data
so more and more references will be included with time.
Where a reference describes a PDB structure, the PDB
codes are
shown on the right.
|
|


 Links
Links






































