 |
PDBsum entry 2amp

|
|
|
|
|
 |
Contents |
 |
|
|
|
|
|
|
|
|
|
|
|
|
* Residue conservation analysis
|
|
|
|
|
|
|
|
|
|
|
Enzyme class 2:
|
|
E.C.2.1.1.57
- methyltransferase cap1.
|
|
|
|
|
|
|
Reaction:
|
|
a 5'-end (N(7)-methyl 5'-triphosphoguanosine)-ribonucleoside in mRNA + S-adenosyl-L-methionine = a 5'-end (N(7)-methyl 5'-triphosphoguanosine)- (2'-O-methyl-ribonucleoside) in mRNA + S-adenosyl-L-homocysteine + H+
|
|
|
|
|
|
5'-end (N(7)-methyl 5'-triphosphoguanosine)-ribonucleoside in mRNA
|
+
|
 S-adenosyl-L-methionine
S-adenosyl-L-methionine
|
=
|
5'-end (N(7)-methyl 5'-triphosphoguanosine)- (2'-O-methyl-ribonucleoside) in mRNA
|
+
|
 S-adenosyl-L-homocysteine
S-adenosyl-L-homocysteine
|
+
|
H(+)
|
|
|
|
|
|
|
|
|
|
Enzyme class 3:
|
|
E.C.2.7.7.48
- RNA-directed Rna polymerase.
|
|
|
|
|
|
|
Reaction:
|
|
RNA(n) + a ribonucleoside 5'-triphosphate = RNA(n+1) + diphosphate
|
|
|
|
|
|
 RNA(n)
RNA(n)
|
+
|
ribonucleoside 5'-triphosphate
|
=
|
RNA(n+1)
|
+
|
 diphosphate
diphosphate
|
|
|
|
|
|
|
|
|
|
Enzyme class 4:
|
|
E.C.2.7.7.50
- mRNA guanylyltransferase.
|
|
|
|
|
|
|
Reaction:
|
|
a 5'-end diphospho-ribonucleoside in mRNA + GTP + H+ = a 5'-end (5'-triphosphoguanosine)-ribonucleoside in mRNA + diphosphate
|
|
|
|
|
|
5'-end diphospho-ribonucleoside in mRNA
|
+
|
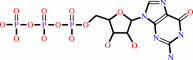 GTP
GTP
|
+
|
H(+)
|
=
|
5'-end (5'-triphosphoguanosine)-ribonucleoside in mRNA
|
+
|
diphosphate
|
|
|
|
|
|
|
|
|
|
Enzyme class 5:
|
|
E.C.3.1.13.-
- ?????
|
|
|
|
|
|
|
Enzyme class 6:
|
|
E.C.3.4.19.12
- ubiquitinyl hydrolase 1.
|
|
|
|
|
|
|
Reaction:
|
|
Thiol-dependent hydrolysis of ester, thiolester, amide, peptide and isopeptide bonds formed by the C-terminal Gly of ubiquitin (a 76-residue protein attached to proteins as an intracellular targeting signal).
|
|
|
|
|
|
Enzyme class 7:
|
|
E.C.3.4.22.-
- ?????
|
|
|
|
|
|
|
Enzyme class 8:
|
|
E.C.3.6.4.12
- Dna helicase.
|
|
|
|
|
|
|
Reaction:
|
|
ATP + H2O = ADP + phosphate + H+
|
|
|
|
|
|
 ATP
ATP
|
+
|
H2O
|
=
|
 ADP
ADP
|
+
|
 phosphate
phosphate
|
+
|
H(+)
|
|
|
|
|
|
|
|
|
|
Enzyme class 9:
|
|
E.C.3.6.4.13
- Rna helicase.
|
|
|
|
|
|
|
Reaction:
|
|
ATP + H2O = ADP + phosphate + H+
|
|
|
|
|
|
ATP
|
+
|
H2O
|
=
|
ADP
|
+
|
phosphate
|
+
|
H(+)
|
|
|
|
|
|
|
|
|
|
Enzyme class 10:
|
|
E.C.4.6.1.-
- ?????
|
|
|
|
|
|
|
|
|
|
Note, where more than one E.C. class is given (as above), each may
correspond to a different protein domain or, in the case of polyprotein
precursors, to a different mature protein.
|
|
|
|
Molecule diagrams generated from .mol files obtained from the
KEGG ftp site
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
| |
|
|
| |
|
|
Plos Biol
3:e324-334
(2005)
|
|
PubMed id:
|
|
|
|
|
|
| |
|
Design of wide-spectrum inhibitors targeting coronavirus main proteases.
|
|
H.Yang,
W.Xie,
X.Xue,
K.Yang,
J.Ma,
W.Liang,
Q.Zhao,
Z.Zhou,
D.Pei,
J.Ziebuhr,
R.Hilgenfeld,
K.Y.Yuen,
L.Wong,
G.Gao,
S.Chen,
Z.Chen,
D.Ma,
M.Bartlam,
Z.Rao.
|
|
|
|
|
| |
ABSTRACT
|
|
|
|
| |
|
|
The genus Coronavirus contains about 25 species of coronaviruses (CoVs), which
are important pathogens causing highly prevalent diseases and often severe or
fatal in humans and animals. No licensed specific drugs are available to prevent
their infection. Different host receptors for cellular entry, poorly conserved
structural proteins (antigens), and the high mutation and recombination rates of
CoVs pose a significant problem in the development of wide-spectrum anti-CoV
drugs and vaccines. CoV main proteases (M(pro)s), which are key enzymes in viral
gene expression and replication, were revealed to share a highly conservative
substrate-recognition pocket by comparison of four crystal structures and a
homology model representing all three genetic clusters of the genus Coronavirus.
This conclusion was further supported by enzyme activity assays. Mechanism-based
irreversible inhibitors were designed, based on this conserved structural
region, and a uniform inhibition mechanism was elucidated from the structures of
Mpro-inhibitor complexes from severe acute respiratory syndrome-CoV and porcine
transmissible gastroenteritis virus. A structure-assisted optimization program
has yielded compounds with fast in vitro inactivation of multiple CoV M(pro)s,
potent antiviral activity, and extremely low cellular toxicity in cell-based
assays. Further modification could rapidly lead to the discovery of a single
agent with clinical potential against existing and possible future emerging
CoV-related diseases.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
 |
 |
|
Literature references that cite this PDB file's key reference
|
|
|
| |
PubMed id
|
|
Reference
|
|
|
|
|
|
H.M.Wang,
and
P.H.Liang
(2010).
Picornaviral 3C protease inhibitors and the dual 3C protease/coronaviral 3C-like protease inhibitors.
|
| |
Expert Opin Ther Pat,
20,
59-71.
|
|
|
|
|
|
|
S.C.Cheng,
G.G.Chang,
and
C.Y.Chou
(2010).
Mutation of Glu-166 blocks the substrate-induced dimerization of SARS coronavirus main protease.
|
| |
Biophys J,
98,
1327-1336.
|
|
|
|
|
|
|
S.Fang,
H.Shen,
J.Wang,
F.P.Tay,
and
D.X.Liu
(2010).
Functional and genetic studies of the substrate specificity of coronavirus infectious bronchitis virus 3C-like proteinase.
|
| |
J Virol,
84,
7325-7336.
|
|
|
|
|
|
|
C.L.Wohlford-Lenane,
D.K.Meyerholz,
S.Perlman,
H.Zhou,
D.Tran,
M.E.Selsted,
and
P.B.McCray
(2009).
Rhesus theta-defensin prevents death in a mouse model of severe acute respiratory syndrome coronavirus pulmonary disease.
|
| |
J Virol,
83,
11385-11390.
|
|
|
|
|
|
|
K.Fiehler,
M.Burke,
S.Bien,
B.Röder,
and
F.Rösler
(2009).
The human dorsal action control system develops in the absence of vision.
|
| |
Cereb Cortex,
19,
1.
|
|
|
|
|
|
|
N.Zhong,
S.Zhang,
F.Xue,
X.Kang,
P.Zou,
J.Chen,
C.Liang,
Z.Rao,
C.Jin,
Z.Lou,
and
B.Xia
(2009).
C-terminal domain of SARS-CoV main protease can form a 3D domain-swapped dimer.
|
| |
Protein Sci,
18,
839-844.
|
|
|
PDB codes:
|
|
|
|
|
|
|
|
Y.Ma,
Y.Feng,
D.Liu,
and
G.F.Gao
(2009).
Avian influenza virus, Streptococcus suis serotype 2, severe acute respiratory syndrome-coronavirus and beyond: molecular epidemiology, ecology and the situation in China.
|
| |
Philos Trans R Soc Lond B Biol Sci,
364,
2725-2737.
|
|
|
|
|
|
|
C.Niu,
J.Yin,
J.Zhang,
J.C.Vederas,
and
M.N.James
(2008).
Molecular docking identifies the binding of 3-chloropyridine moieties specifically to the S1 pocket of SARS-CoV Mpro.
|
| |
Bioorg Med Chem,
16,
293-302.
|
|
|
|
|
|
|
J.Weigelt,
L.D.McBroom-Cerajewski,
M.Schapira,
Y.Zhao,
C.H.Arrowsmith,
and
C.H.Arrowmsmith
(2008).
Structural genomics and drug discovery: all in the family.
|
| |
Curr Opin Chem Biol,
12,
32-39.
|
|
|
|
|
|
|
M.Bartlam,
X.Xue,
and
Z.Rao
(2008).
The search for a structural basis for therapeutic intervention against the SARS coronavirus.
|
| |
Acta Crystallogr A,
64,
204-213.
|
|
|
|
|
|
|
N.Zhong,
S.Zhang,
P.Zou,
J.Chen,
X.Kang,
Z.Li,
C.Liang,
C.Jin,
and
B.Xia
(2008).
Without its N-finger, the main protease of severe acute respiratory syndrome coronavirus can form a novel dimer through its C-terminal domain.
|
| |
J Virol,
82,
4227-4234.
|
|
|
|
|
|
|
Q.Zhao,
S.Li,
F.Xue,
Y.Zou,
C.Chen,
M.Bartlam,
and
Z.Rao
(2008).
Structure of the main protease from a global infectious human coronavirus, HCoV-HKU1.
|
| |
J Virol,
82,
8647-8655.
|
|
|
PDB code:
|
|
|
|
|
|
|
|
U.Bacha,
J.Barrila,
S.B.Gabelli,
Y.Kiso,
L.Mario Amzel,
and
E.Freire
(2008).
Development of broad-spectrum halomethyl ketone inhibitors against coronavirus main protease 3CL(pro).
|
| |
Chem Biol Drug Des,
72,
34-49.
|
|
|
PDB code:
|
|
|
|
|
|
|
|
X.Xue,
H.Yu,
H.Yang,
F.Xue,
Z.Wu,
W.Shen,
J.Li,
Z.Zhou,
Y.Ding,
Q.Zhao,
X.C.Zhang,
M.Liao,
M.Bartlam,
and
Z.Rao
(2008).
Structures of two coronavirus main proteases: implications for substrate binding and antiviral drug design.
|
| |
J Virol,
82,
2515-2527.
|
|
|
PDB codes:
|
|
|
|
|
|
|
|
D.Plewczynski,
M.Hoffmann,
M.von Grotthuss,
K.Ginalski,
and
L.Rychewski
(2007).
In silico prediction of SARS protease inhibitors by virtual high throughput screening.
|
| |
Chem Biol Drug Des,
69,
269-279.
|
|
|
|
|
|
|
J.Li,
W.Shen,
M.Liao,
and
M.Bartlam
(2007).
Preliminary crystallographic analysis of avian infectious bronchitis virus main protease.
|
| |
Acta Crystallogr Sect F Struct Biol Cryst Commun,
63,
24-26.
|
|
|
|
|
|
|
K.Pyrc,
B.Berkhout,
and
L.van der Hoek
(2007).
The novel human coronaviruses NL63 and HKU1.
|
| |
J Virol,
81,
3051-3057.
|
|
|
|
|
|
|
M.Bartlam,
Y.Xu,
and
Z.Rao
(2007).
Structural proteomics of the SARS coronavirus: a model response to emerging infectious diseases.
|
| |
J Struct Funct Genomics,
8,
85-97.
|
|
|
|
|
|
|
Y.M.Shao,
W.B.Yang,
H.P.Peng,
M.F.Hsu,
K.C.Tsai,
T.H.Kuo,
A.H.Wang,
P.H.Liang,
C.H.Lin,
A.S.Yang,
and
C.H.Wong
(2007).
Structure-based design and synthesis of highly potent SARS-CoV 3CL protease inhibitors.
|
| |
Chembiochem,
8,
1654-1657.
|
|
|
|
|
|
|
Z.Chen,
Y.Wang,
K.Ratia,
A.D.Mesecar,
K.D.Wilkinson,
and
S.C.Baker
(2007).
Proteolytic processing and deubiquitinating activity of papain-like proteases of human coronavirus NL63.
|
| |
J Virol,
81,
6007-6018.
|
|
|
|
|
|
|
Z.Rao
(2007).
History of protein crystallography in China.
|
| |
Philos Trans R Soc Lond B Biol Sci,
362,
1035-1042.
|
|
|
|
|
|
|
A.Mandavilli
(2006).
China: open season.
|
| |
Nature,
439,
382-383.
|
|
|
|
|
|
|
D.Su,
Z.Lou,
F.Sun,
Y.Zhai,
H.Yang,
R.Zhang,
A.Joachimiak,
X.C.Zhang,
M.Bartlam,
and
Z.Rao
(2006).
Dodecamer structure of severe acute respiratory syndrome coronavirus nonstructural protein nsp10.
|
| |
J Virol,
80,
7902-7908.
|
|
|
PDB codes:
|
|
|
|
|
|
|
|
H.Chen,
P.Wei,
C.Huang,
L.Tan,
Y.Liu,
and
L.Lai
(2006).
Only one protomer is active in the dimer of SARS 3C-like proteinase.
|
| |
J Biol Chem,
281,
13894-13898.
|
|
|
|
|
|
|
J.R.Mesters,
J.Tan,
and
R.Hilgenfeld
(2006).
Viral enzymes.
|
| |
Curr Opin Struct Biol,
16,
776-786.
|
|
|
|
|
|
|
J.Shi,
and
J.Song
(2006).
The catalysis of the SARS 3C-like protease is under extensive regulation by its extra domain.
|
| |
FEBS J,
273,
1035-1045.
|
|
|
|
|
|
|
L.van der Hoek,
K.Pyrc,
and
B.Berkhout
(2006).
Human coronavirus NL63, a new respiratory virus.
|
| |
FEMS Microbiol Rev,
30,
760-773.
|
|
|
|
|
|
|
S.I.Al-Gharabli,
S.T.Shah,
S.Weik,
M.F.Schmidt,
J.R.Mesters,
D.Kuhn,
G.Klebe,
R.Hilgenfeld,
and
J.Rademann
(2006).
An efficient method for the synthesis of peptide aldehyde libraries employed in the discovery of reversible SARS coronavirus main protease (SARS-CoV Mpro) inhibitors.
|
| |
Chembiochem,
7,
1048-1055.
|
|
|
|
|
|
|
X.Xu,
Y.Zhai,
F.Sun,
Z.Lou,
D.Su,
Y.Xu,
R.Zhang,
A.Joachimiak,
X.C.Zhang,
M.Bartlam,
and
Z.Rao
(2006).
New antiviral target revealed by the hexameric structure of mouse hepatitis virus nonstructural protein nsp15.
|
| |
J Virol,
80,
7909-7917.
|
|
|
PDB codes:
|
|
|
|
|
|
|
The most recent references are shown first.
Citation data come partly from CiteXplore and partly
from an automated harvesting procedure. Note that this is likely to be
only a partial list as not all journals are covered by
either method. However, we are continually building up the citation data
so more and more references will be included with time.
Where a reference describes a PDB structure, the PDB
codes are
shown on the right.
|
|


 Links
Links




































