 |
PDBsum entry 1lvo

|
|
|
|
|
 |
Contents |
 |
|
|
|
|
|
|
|
|
|
|
|
|
* Residue conservation analysis
|
|
|
|
|
|
PDB id:
|
|
|
|
| Name: |
|
Hydrolase
|
|
|
Title:
|
|
Structure of coronavirus main proteinase reveals combination of a chymotrypsin fold with an extra alpha-helical domain
|
|
Structure:
|
|
Replicase, hydrolase domain. Chain: a, b, c, d, e, f. Fragment: residues 2879-3180. Engineered: yes
|
|
Source:
|
|
Transmissible gastroenteritis virus. Organism_taxid: 11149. Gene: orf1a. Expressed in: escherichia coli. Expression_system_taxid: 562.
|
|
Biol. unit:
|
|
 Dimer (from
)
Dimer (from
)
|
|
Resolution:
|
|
|
1.96Å
|
R-factor:
|
0.210
|
R-free:
|
0.256
|
|
|
Authors:
|
|
K.Anand,G.J.Palm,J.R.Mesters,S.G.Siddell,J.Ziebuhr,R.Hilgenfeld
|
Key ref:

|
|
K.Anand
et al.
(2002).
Structure of coronavirus main proteinase reveals combination of a chymotrypsin fold with an extra alpha-helical domain.
EMBO J,
21,
3213-3224.
PubMed id:
DOI:

|
|
|
Date:
|
|
|
29-May-02
|
Release date:
|
17-Jul-02
|
|
|
|
|
|
|
PROCHECK
|

|
|
|
|
|
Headers
|
|
|
|
References
|
|
|
|
|
|

|
|
P0C6Y5
(R1AB_CVPPU) -
Replicase polyprotein 1ab from Porcine transmissible gastroenteritis coronavirus (strain Purdue)
|
|
|
|
Seq:
Struc:
|
|
|
|
6684 a.a.
299 a.a.
|
|

|

|
|
|
|
|
|
|
|
|
|
|
|
Key: |
 |
PfamA domain |
|
|
 |
Secondary structure |
|
 |
CATH domain |
|
|
|
|
|
|
|
|
|
|
|
|
|
Enzyme class 2:
|
|
E.C.2.1.1.57
- methyltransferase cap1.
|
|
|
|
|
|
|
Reaction:
|
|
a 5'-end (N(7)-methyl 5'-triphosphoguanosine)-ribonucleoside in mRNA + S-adenosyl-L-methionine = a 5'-end (N(7)-methyl 5'-triphosphoguanosine)- (2'-O-methyl-ribonucleoside) in mRNA + S-adenosyl-L-homocysteine + H+
|
|
|
|
|
|
5'-end (N(7)-methyl 5'-triphosphoguanosine)-ribonucleoside in mRNA
|
+
|
 S-adenosyl-L-methionine
S-adenosyl-L-methionine
|
=
|
5'-end (N(7)-methyl 5'-triphosphoguanosine)- (2'-O-methyl-ribonucleoside) in mRNA
|
+
|
 S-adenosyl-L-homocysteine
S-adenosyl-L-homocysteine
|
+
|
H(+)
|
|
|
|
|
|
|
|
|
|
Enzyme class 3:
|
|
E.C.2.7.7.48
- RNA-directed Rna polymerase.
|
|
|
|
|
|
|
Reaction:
|
|
RNA(n) + a ribonucleoside 5'-triphosphate = RNA(n+1) + diphosphate
|
|
|
|
|
|
 RNA(n)
RNA(n)
|
+
|
ribonucleoside 5'-triphosphate
|
=
|
RNA(n+1)
|
+
|
 diphosphate
diphosphate
|
|
|
|
|
|
|
|
|
|
Enzyme class 4:
|
|
E.C.2.7.7.50
- mRNA guanylyltransferase.
|
|
|
|
|
|
|
Reaction:
|
|
a 5'-end diphospho-ribonucleoside in mRNA + GTP + H+ = a 5'-end (5'-triphosphoguanosine)-ribonucleoside in mRNA + diphosphate
|
|
|
|
|
|
5'-end diphospho-ribonucleoside in mRNA
|
+
|
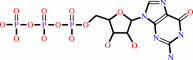 GTP
GTP
|
+
|
H(+)
|
=
|
5'-end (5'-triphosphoguanosine)-ribonucleoside in mRNA
|
+
|
diphosphate
|
|
|
|
|
|
|
|
|
|
Enzyme class 5:
|
|
E.C.3.1.13.-
- ?????
|
|
|
|
|
|
|
Enzyme class 6:
|
|
E.C.3.4.19.12
- ubiquitinyl hydrolase 1.
|
|
|
|
|
|
|
Reaction:
|
|
Thiol-dependent hydrolysis of ester, thiolester, amide, peptide and isopeptide bonds formed by the C-terminal Gly of ubiquitin (a 76-residue protein attached to proteins as an intracellular targeting signal).
|
|
|
|
|
|
Enzyme class 7:
|
|
E.C.3.4.22.-
- ?????
|
|
|
|
|
|
|
Enzyme class 8:
|
|
E.C.3.6.4.12
- Dna helicase.
|
|
|
|
|
|
|
Reaction:
|
|
ATP + H2O = ADP + phosphate + H+
|
|
|
|
|
|
 ATP
ATP
|
+
|
H2O
|
=
|
 ADP
ADP
|
+
|
 phosphate
phosphate
|
+
|
H(+)
|
|
|
|
|
|
|
|
|
|
Enzyme class 9:
|
|
E.C.3.6.4.13
- Rna helicase.
|
|
|
|
|
|
|
Reaction:
|
|
ATP + H2O = ADP + phosphate + H+
|
|
|
|
|
|
ATP
|
+
|
H2O
|
=
|
ADP
|
+
|
phosphate
|
+
|
H(+)
|
|
|
|
|
|
|
|
|
|
Enzyme class 10:
|
|
E.C.4.6.1.-
- ?????
|
|
|
|
|
|
|
|
|
|
Note, where more than one E.C. class is given (as above), each may
correspond to a different protein domain or, in the case of polyprotein
precursors, to a different mature protein.
|
|
|
|
Molecule diagrams generated from .mol files obtained from the
KEGG ftp site
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
| |
|
|
| |
|
DOI no:
|
EMBO J
21:3213-3224
(2002)
|
|
PubMed id:
|
|
|
|
|
|
| |
|
Structure of coronavirus main proteinase reveals combination of a chymotrypsin fold with an extra alpha-helical domain.
|
|
K.Anand,
G.J.Palm,
J.R.Mesters,
S.G.Siddell,
J.Ziebuhr,
R.Hilgenfeld.
|
|
|
|
|
| |
ABSTRACT
|
|
|
|
| |
|
|
The key enzyme in coronavirus polyprotein processing is the viral main
proteinase, M(pro), a protein with extremely low sequence similarity to other
viral and cellular proteinases. Here, the crystal structure of the 33.1 kDa
transmissible gastroenteritis (corona)virus M(pro) is reported. The structure
was refined to 1.96 A resolution and revealed three dimers in the asymmetric
unit. The mutual arrangement of the protomers in each of the dimers suggests
that M(pro) self-processing occurs in trans. The active site, comprised of
Cys144 and His41, is part of a chymotrypsin-like fold that is connected by a 16
residue loop to an extra domain featuring a novel alpha-helical fold. Molecular
modelling and mutagenesis data implicate the loop in substrate binding and
elucidate S1 and S2 subsites suitable to accommodate the side chains of the P1
glutamine and P2 leucine residues of M(pro) substrates. Interactions involving
the N-terminus and the alpha-helical domain stabilize the loop in the
orientation required for trans-cleavage activity. The study illustrates that RNA
viruses have evolved unprecedented variations of the classical chymotrypsin fold.
|
|
|
|
|
|
| |
Selected figure(s)
|
|
|
|
| |
|
|

|
|

|
|
Figure 7.
Figure 7 Stereo diagram of a P5−P1 substrate
(Asn−Ser−Thr−Leu−Gln, red; corresponding to the TGEV
M^pro N-terminal autoprocessing site) modelled into the active
site cleft of the TGEV M^pro. Hydrogen bonds are depicted by
dotted lines.
|
|
Figure 8.
Figure 8 Intra- and intermolecular contacts of the TGEV M^pro
N-terminus. (A) MOLSCRIPT stereo representation of a TGEV M^pro
dimer. Molecule A is coloured from blue at the N-terminus, via
green (domain II), to red (C-terminus), while molecule B is
shown in grey. The catalytic Cys144 and His41 residues are
labelled in both monomers. (B) Detailed view of the interactions
made by the N-terminal segment (blue) and domains II/III of
monomer A as well as domains II/III of monomer B. Residues
critically involved in these interactions are designated by the
single-letter code and shown in ball-and-stick representation
(see text for details). The N- and C-termini of molecule A are
indicated.
|
|
|
|
|
|
| |
The above figures are
reprinted
from an Open Access publication published by Macmillan Publishers Ltd:
EMBO J
(2002,
21,
3213-3224)
copyright 2002.
|
|
| |
Figures were
selected
by an automated process.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
 |
 |
|
Literature references that cite this PDB file's key reference
|
|
|
| |
PubMed id
|
|
Reference
|
|
|
|
|
|
J.Shi,
N.Han,
L.Lim,
S.Lua,
J.Sivaraman,
L.Wang,
Y.Mu,
and
J.Song
(2011).
Dynamically-driven inactivation of the catalytic machinery of the SARS 3C-like protease by the N214A mutation on the extra domain.
|
| |
PLoS Comput Biol,
7,
e1001084.
|
|
|
|
|
|
|
M.Y.Tsai,
W.H.Chang,
J.Y.Liang,
L.L.Lin,
G.G.Chang,
and
H.P.Chang
(2010).
Essential covalent linkage between the chymotrypsin-like domain and the extra domain of the SARS-CoV main protease.
|
| |
J Biochem,
148,
349-358.
|
|
|
|
|
|
|
R.N.Kostoff
(2010).
The highly cited SARS research literature.
|
| |
Crit Rev Microbiol,
36,
299-317.
|
|
|
|
|
|
|
S.C.Cheng,
G.G.Chang,
and
C.Y.Chou
(2010).
Mutation of Glu-166 blocks the substrate-induced dimerization of SARS coronavirus main protease.
|
| |
Biophys J,
98,
1327-1336.
|
|
|
|
|
|
|
S.Fang,
H.Shen,
J.Wang,
F.P.Tay,
and
D.X.Liu
(2010).
Functional and genetic studies of the substrate specificity of coronavirus infectious bronchitis virus 3C-like proteinase.
|
| |
J Virol,
84,
7325-7336.
|
|
|
|
|
|
|
N.Zhong,
S.Zhang,
F.Xue,
X.Kang,
P.Zou,
J.Chen,
C.Liang,
Z.Rao,
C.Jin,
Z.Lou,
and
B.Xia
(2009).
C-terminal domain of SARS-CoV main protease can form a 3D domain-swapped dimer.
|
| |
Protein Sci,
18,
839-844.
|
|
|
PDB codes:
|
|
|
|
|
|
|
|
R.Caliandro,
B.Carrozzini,
G.L.Cascarano,
C.Giacovazzo,
A.M.Mazzone,
and
D.Siliqi
(2009).
EDM-DEDM and protein crystal structure solution.
|
| |
Acta Crystallogr D Biol Crystallogr,
65,
477-484.
|
|
|
|
|
|
|
Y.Xu,
L.Cong,
C.Chen,
L.Wei,
Q.Zhao,
X.Xu,
Y.Ma,
M.Bartlam,
and
Z.Rao
(2009).
Crystal structures of two coronavirus ADP-ribose-1''-monophosphatases and their complexes with ADP-Ribose: a systematic structural analysis of the viral ADRP domain.
|
| |
J Virol,
83,
1083-1092.
|
|
|
PDB codes:
|
|
|
|
|
|
|
|
C.Niu,
J.Yin,
J.Zhang,
J.C.Vederas,
and
M.N.James
(2008).
Molecular docking identifies the binding of 3-chloropyridine moieties specifically to the S1 pocket of SARS-CoV Mpro.
|
| |
Bioorg Med Chem,
16,
293-302.
|
|
|
|
|
|
|
I.Robel,
J.Gebhardt,
J.R.Mesters,
A.Gorbalenya,
B.Coutard,
B.Canard,
R.Hilgenfeld,
and
J.Rohayem
(2008).
Functional characterization of the cleavage specificity of the sapovirus chymotrypsin-like protease.
|
| |
J Virol,
82,
8085-8093.
|
|
|
|
|
|
|
J.S.Sparks,
E.F.Donaldson,
X.Lu,
R.S.Baric,
and
M.R.Denison
(2008).
A novel mutation in murine hepatitis virus nsp5, the viral 3C-like proteinase, causes temperature-sensitive defects in viral growth and protein processing.
|
| |
J Virol,
82,
5999-6008.
|
|
|
|
|
|
|
J.Shi,
J.Sivaraman,
and
J.Song
(2008).
Mechanism for controlling the dimer-monomer switch and coupling dimerization to catalysis of the severe acute respiratory syndrome coronavirus 3C-like protease.
|
| |
J Virol,
82,
4620-4629.
|
|
|
PDB code:
|
|
|
|
|
|
|
|
M.Bartlam,
X.Xue,
and
Z.Rao
(2008).
The search for a structural basis for therapeutic intervention against the SARS coronavirus.
|
| |
Acta Crystallogr A,
64,
204-213.
|
|
|
|
|
|
|
N.Zhong,
S.Zhang,
P.Zou,
J.Chen,
X.Kang,
Z.Li,
C.Liang,
C.Jin,
and
B.Xia
(2008).
Without its N-finger, the main protease of severe acute respiratory syndrome coronavirus can form a novel dimer through its C-terminal domain.
|
| |
J Virol,
82,
4227-4234.
|
|
|
|
|
|
|
Q.Zhao,
S.Li,
F.Xue,
Y.Zou,
C.Chen,
M.Bartlam,
and
Z.Rao
(2008).
Structure of the main protease from a global infectious human coronavirus, HCoV-HKU1.
|
| |
J Virol,
82,
8647-8655.
|
|
|
PDB code:
|
|
|
|
|
|
|
|
U.Bacha,
J.Barrila,
S.B.Gabelli,
Y.Kiso,
L.Mario Amzel,
and
E.Freire
(2008).
Development of broad-spectrum halomethyl ketone inhibitors against coronavirus main protease 3CL(pro).
|
| |
Chem Biol Drug Des,
72,
34-49.
|
|
|
PDB code:
|
|
|
|
|
|
|
|
X.Xue,
H.Yu,
H.Yang,
F.Xue,
Z.Wu,
W.Shen,
J.Li,
Z.Zhou,
Y.Ding,
Q.Zhao,
X.C.Zhang,
M.Liao,
M.Bartlam,
and
Z.Rao
(2008).
Structures of two coronavirus main proteases: implications for substrate binding and antiviral drug design.
|
| |
J Virol,
82,
2515-2527.
|
|
|
PDB codes:
|
|
|
|
|
|
|
|
Y.Someya,
N.Takeda,
and
T.Wakita
(2008).
Saturation Mutagenesis reveals that GLU54 of Norovirus 3C-like Protease is not Essential for the Proteolytic Activity.
|
| |
J Biochem,
144,
771-780.
|
|
|
|
|
|
|
A.K.Ghosh,
K.Xi,
M.E.Johnson,
S.C.Baker,
and
A.D.Mesecar
(2007).
Progress in Anti-SARS Coronavirus Chemistry, Biology and Chemotherapy.
|
| |
Annu Rep Med Chem,
41,
183-196.
|
|
|
|
|
|
|
C.W.Yang,
Y.N.Yang,
P.H.Liang,
C.M.Chen,
W.L.Chen,
H.Y.Chang,
Y.S.Chao,
and
S.J.Lee
(2007).
Novel small-molecule inhibitors of transmissible gastroenteritis virus.
|
| |
Antimicrob Agents Chemother,
51,
3924-3931.
|
|
|
|
|
|
|
E.F.Donaldson,
R.L.Graham,
A.C.Sims,
M.R.Denison,
and
R.S.Baric
(2007).
Analysis of murine hepatitis virus strain A59 temperature-sensitive mutant TS-LA6 suggests that nsp10 plays a critical role in polyprotein processing.
|
| |
J Virol,
81,
7086-7098.
|
|
|
|
|
|
|
G.Fear,
S.Komarnytsky,
and
I.Raskin
(2007).
Protease inhibitors and their peptidomimetic derivatives as potential drugs.
|
| |
Pharmacol Ther,
113,
354-368.
|
|
|
|
|
|
|
H.P.Chang,
C.Y.Chou,
and
G.G.Chang
(2007).
Reversible unfolding of the severe acute respiratory syndrome coronavirus main protease in guanidinium chloride.
|
| |
Biophys J,
92,
1374-1383.
|
|
|
|
|
|
|
J.Li,
W.Shen,
M.Liao,
and
M.Bartlam
(2007).
Preliminary crystallographic analysis of avian infectious bronchitis virus main protease.
|
| |
Acta Crystallogr Sect F Struct Biol Cryst Commun,
63,
24-26.
|
|
|
|
|
|
|
J.Ziebuhr,
B.Schelle,
N.Karl,
E.Minskaia,
S.Bayer,
S.G.Siddell,
A.E.Gorbalenya,
and
V.Thiel
(2007).
Human coronavirus 229E papain-like proteases have overlapping specificities but distinct functions in viral replication.
|
| |
J Virol,
81,
3922-3932.
|
|
|
|
|
|
|
K.Zheng,
G.Ma,
J.Zhou,
M.Zen,
W.Zhao,
Y.Jiang,
Q.Yu,
and
J.Feng
(2007).
Insight into the activity of SARS main protease: Molecular dynamics study of dimeric and monomeric form of enzyme.
|
| |
Proteins,
66,
467-479.
|
|
|
|
|
|
|
M.Bartlam,
Y.Xu,
and
Z.Rao
(2007).
Structural proteomics of the SARS coronavirus: a model response to emerging infectious diseases.
|
| |
J Struct Funct Genomics,
8,
85-97.
|
|
|
|
|
|
|
S.Curry,
N.Roqué-Rosell,
P.A.Zunszain,
and
R.J.Leatherbarrow
(2007).
Foot-and-mouth disease virus 3C protease: recent structural and functional insights into an antiviral target.
|
| |
Int J Biochem Cell Biol,
39,
1-6.
|
|
|
|
|
|
|
S.G.Sawicki,
D.L.Sawicki,
and
S.G.Siddell
(2007).
A contemporary view of coronavirus transcription.
|
| |
J Virol,
81,
20-29.
|
|
|
|
|
|
|
X.Tian,
Y.Feng,
T.Zhao,
H.Peng,
J.Yan,
J.Qi,
F.Jiang,
K.Tian,
and
F.Gao
(2007).
Molecular cloning, expression, purification and crystallographic analysis of PRRSV 3CL protease.
|
| |
Acta Crystallogr Sect F Struct Biol Cryst Commun,
63,
720-722.
|
|
|
|
|
|
|
A.E.Gorbalenya,
L.Enjuanes,
J.Ziebuhr,
and
E.J.Snijder
(2006).
Nidovirales: evolving the largest RNA virus genome.
|
| |
Virus Res,
117,
17-37.
|
|
|
|
|
|
|
D.van Aken,
E.J.Snijder,
and
A.E.Gorbalenya
(2006).
Mutagenesis analysis of the nsp4 main proteinase reveals determinants of arterivirus replicase polyprotein autoprocessing.
|
| |
J Virol,
80,
3428-3437.
|
|
|
|
|
|
|
H.Chen,
P.Wei,
C.Huang,
L.Tan,
Y.Liu,
and
L.Lai
(2006).
Only one protomer is active in the dimer of SARS 3C-like proteinase.
|
| |
J Biol Chem,
281,
13894-13898.
|
|
|
|
|
|
|
H.Schütze,
R.Ulferts,
B.Schelle,
S.Bayer,
H.Granzow,
B.Hoffmann,
T.C.Mettenleiter,
and
J.Ziebuhr
(2006).
Characterization of White bream virus reveals a novel genetic cluster of nidoviruses.
|
| |
J Virol,
80,
11598-11609.
|
|
|
|
|
|
|
J.Barrila,
U.Bacha,
and
E.Freire
(2006).
Long-range cooperative interactions modulate dimerization in SARS 3CLpro.
|
| |
Biochemistry,
45,
14908-14916.
|
|
|
|
|
|
|
J.R.Mesters,
J.Tan,
and
R.Hilgenfeld
(2006).
Viral enzymes.
|
| |
Curr Opin Struct Biol,
16,
776-786.
|
|
|
|
|
|
|
J.Shi,
and
J.Song
(2006).
The catalysis of the SARS 3C-like protease is under extensive regulation by its extra domain.
|
| |
FEBS J,
273,
1035-1045.
|
|
|
|
|
|
|
P.Hamill,
D.Hudson,
R.Y.Kao,
P.Chow,
M.Raj,
H.Xu,
M.J.Richer,
and
F.Jean
(2006).
Development of a red-shifted fluorescence-based assay for SARS-coronavirus 3CL protease: identification of a novel class of anti-SARS agents from the tropical marine sponge Axinella corrugata.
|
| |
Biol Chem,
387,
1063-1074.
|
|
|
|
|
|
|
S.L.Smits,
E.J.Snijder,
and
R.J.de Groot
(2006).
Characterization of a torovirus main proteinase.
|
| |
J Virol,
80,
4157-4167.
|
|
|
|
|
|
|
A.Putics,
W.Filipowicz,
J.Hall,
A.E.Gorbalenya,
and
J.Ziebuhr
(2005).
ADP-ribose-1"-monophosphatase: a conserved coronavirus enzyme that is dispensable for viral replication in tissue culture.
|
| |
J Virol,
79,
12721-12731.
|
|
|
|
|
|
|
D.A.Groneberg,
R.Hilgenfeld,
and
P.Zabel
(2005).
Molecular mechanisms of severe acute respiratory syndrome (SARS).
|
| |
Respir Res,
6,
8.
|
|
|
|
|
|
|
H.Yang,
W.Xie,
X.Xue,
K.Yang,
J.Ma,
W.Liang,
Q.Zhao,
Z.Zhou,
D.Pei,
J.Ziebuhr,
R.Hilgenfeld,
K.Y.Yuen,
L.Wong,
G.Gao,
S.Chen,
Z.Chen,
D.Ma,
M.Bartlam,
and
Z.Rao
(2005).
Design of wide-spectrum inhibitors targeting coronavirus main proteases.
|
| |
PLoS Biol,
3,
e324.
|
|
|
PDB codes:
|
|
|
|
|
|
|
|
K.Nakamura,
Y.Someya,
T.Kumasaka,
G.Ueno,
M.Yamamoto,
T.Sato,
N.Takeda,
T.Miyamura,
and
N.Tanaka
(2005).
A norovirus protease structure provides insights into active and substrate binding site integrity.
|
| |
J Virol,
79,
13685-13693.
|
|
|
PDB code:
|
|
|
|
|
|
|
|
M.F.Hsu,
C.J.Kuo,
K.T.Chang,
H.C.Chang,
C.C.Chou,
T.P.Ko,
H.L.Shr,
G.G.Chang,
A.H.Wang,
and
P.H.Liang
(2005).
Mechanism of the maturation process of SARS-CoV 3CL protease.
|
| |
J Biol Chem,
280,
31257-31266.
|
|
|
PDB codes:
|
|
|
|
|
|
|
|
S.Chen,
L.Chen,
J.Tan,
J.Chen,
L.Du,
T.Sun,
J.Shen,
K.Chen,
H.Jiang,
and
X.Shen
(2005).
Severe acute respiratory syndrome coronavirus 3C-like proteinase N terminus is indispensable for proteolytic activity but not for enzyme dimerization. Biochemical and thermodynamic investigation in conjunction with molecular dynamics simulations.
|
| |
J Biol Chem,
280,
164-173.
|
|
|
|
|
|
|
S.Chen,
L.L.Chen,
H.B.Luo,
T.Sun,
J.Chen,
F.Ye,
J.H.Cai,
J.K.Shen,
X.Shen,
and
H.L.Jiang
(2005).
Enzymatic activity characterization of SARS coronavirus 3C-like protease by fluorescence resonance energy transfer technique.
|
| |
Acta Pharmacol Sin,
26,
99.
|
|
|
|
|
|
|
S.G.Sawicki,
D.L.Sawicki,
D.Younker,
Y.Meyer,
V.Thiel,
H.Stokes,
and
S.G.Siddell
(2005).
Functional and genetic analysis of coronavirus replicase-transcriptase proteins.
|
| |
PLoS Pathog,
1,
e39.
|
|
|
|
|
|
|
W.C.Hsu,
H.C.Chang,
C.Y.Chou,
P.J.Tsai,
P.I.Lin,
and
G.G.Chang
(2005).
Critical assessment of important regions in the subunit association and catalytic action of the severe acute respiratory syndrome coronavirus main protease.
|
| |
J Biol Chem,
280,
22741-22748.
|
|
|
|
|
|
|
Y.F.Shan,
and
G.J.Xu
(2005).
Study on substrate specificity at subsites for severe acute respiratory syndrome coronavirus 3CL protease.
|
| |
Acta Biochim Biophys Sin (Shanghai),
37,
807-813.
|
|
|
|
|
|
|
A.Hillisch,
L.F.Pineda,
and
R.Hilgenfeld
(2004).
Utility of homology models in the drug discovery process.
|
| |
Drug Discov Today,
9,
659-669.
|
|
|
|
|
|
|
G.Sutton,
E.Fry,
L.Carter,
S.Sainsbury,
T.Walter,
J.Nettleship,
N.Berrow,
R.Owens,
R.Gilbert,
A.Davidson,
S.Siddell,
L.L.Poon,
J.Diprose,
D.Alderton,
M.Walsh,
J.M.Grimes,
and
D.I.Stuart
(2004).
The nsp9 replicase protein of SARS-coronavirus, structure and functional insights.
|
| |
Structure,
12,
341-353.
|
|
|
PDB code:
|
|
|
|
|
|
|
|
J.Kopp,
and
T.Schwede
(2004).
Automated protein structure homology modeling: a progress report.
|
| |
Pharmacogenomics,
5,
405-416.
|
|
|
|
|
|
|
J.R.St-Jean,
H.Jacomy,
M.Desforges,
A.Vabret,
F.Freymuth,
and
P.J.Talbot
(2004).
Human respiratory coronavirus OC43: genetic stability and neuroinvasion.
|
| |
J Virol,
78,
8824-8834.
|
|
|
|
|
|
|
J.Shi,
Z.Wei,
and
J.Song
(2004).
Dissection study on the severe acute respiratory syndrome 3C-like protease reveals the critical role of the extra domain in dimerization of the enzyme: defining the extra domain as a new target for design of highly specific protease inhibitors.
|
| |
J Biol Chem,
279,
24765-24773.
|
|
|
|
|
|
|
J.Ziebuhr
(2004).
Molecular biology of severe acute respiratory syndrome coronavirus.
|
| |
Curr Opin Microbiol,
7,
412-419.
|
|
|
|
|
|
|
K.Fan,
P.Wei,
Q.Feng,
S.Chen,
C.Huang,
L.Ma,
B.Lai,
J.Pei,
Y.Liu,
J.Chen,
and
L.Lai
(2004).
Biosynthesis, purification, and substrate specificity of severe acute respiratory syndrome coronavirus 3C-like proteinase.
|
| |
J Biol Chem,
279,
1637-1642.
|
|
|
|
|
|
|
M.C.Burla,
B.Carrozzini,
G.L.Cascarano,
C.Giacovazzo,
M.Moustiakimov,
G.Polidori,
and
D.Siliqi
(2004).
MAD phasing: choosing the most informative wavelength combination.
|
| |
Acta Crystallogr D Biol Crystallogr,
60,
1683-1686.
|
|
|
|
|
|
|
M.Parera,
B.Clotet,
and
M.A.Martinez
(2004).
Genetic screen for monitoring severe acute respiratory syndrome coronavirus 3C-like protease.
|
| |
J Virol,
78,
14057-14061.
|
|
|
|
|
|
|
M.Takeda-Shitaka,
H.Nojima,
D.Takaya,
K.Kanou,
M.Iwadate,
and
H.Umeyama
(2004).
Evaluation of homology modeling of the severe acute respiratory syndrome (SARS) coronavirus main protease for structure based drug design.
|
| |
Chem Pharm Bull (Tokyo),
52,
643-645.
|
|
|
|
|
|
|
Y.P.Pang
(2004).
Three-dimensional model of a substrate-bound SARS chymotrypsin-like cysteine proteinase predicted by multiple molecular dynamics simulations: catalytic efficiency regulated by substrate binding.
|
| |
Proteins,
57,
747-757.
|
|
|
PDB codes:
|
|
|
|
|
|
|
|
A.Davidson,
and
S.Siddell
(2003).
Potential for antiviral treatment of severe acute respiratory syndrome.
|
| |
Curr Opin Infect Dis,
16,
565-571.
|
|
|
|
|
|
|
A.Kanjanahaluethai,
D.Jukneliene,
and
S.C.Baker
(2003).
Identification of the murine coronavirus MP1 cleavage site recognized by papain-like proteinase 2.
|
| |
J Virol,
77,
7376-7382.
|
|
|
|
|
|
|
H.Yang,
M.Yang,
Y.Ding,
Y.Liu,
Z.Lou,
Z.Zhou,
L.Sun,
L.Mo,
S.Ye,
H.Pang,
G.F.Gao,
K.Anand,
M.Bartlam,
R.Hilgenfeld,
and
Z.Rao
(2003).
The crystal structures of severe acute respiratory syndrome virus main protease and its complex with an inhibitor.
|
| |
Proc Natl Acad Sci U S A,
100,
13190-13195.
|
|
|
PDB codes:
|
|
|
|
|
|
|
|
J.Ziebuhr,
S.Bayer,
J.A.Cowley,
and
A.E.Gorbalenya
(2003).
The 3C-like proteinase of an invertebrate nidovirus links coronavirus and potyvirus homologs.
|
| |
J Virol,
77,
1415-1426.
|
|
|
|
|
|
|
K.Anand,
J.Ziebuhr,
P.Wadhwani,
J.R.Mesters,
and
R.Hilgenfeld
(2003).
Coronavirus main proteinase (3CLpro) structure: basis for design of anti-SARS drugs.
|
| |
Science,
300,
1763-1767.
|
|
|
PDB codes:
|
|
|
|
|
|
|
|
K.Stadler,
V.Masignani,
M.Eickmann,
S.Becker,
S.Abrignani,
H.D.Klenk,
and
R.Rappuoli
(2003).
SARS--beginning to understand a new virus.
|
| |
Nat Rev Microbiol,
1,
209-218.
|
|
|
|
|
|
|
M.C.Burla,
B.Carrozzini,
G.L.Cascarano,
C.Giacovazzo,
and
G.Polidori
(2003).
SAD or MAD phasing: location of the anomalous scatterers.
|
| |
Acta Crystallogr D Biol Crystallogr,
59,
662-669.
|
|
|
|
|
|
|
I.H.Barrette-Ng,
K.K.Ng,
B.L.Mark,
D.Van Aken,
M.M.Cherney,
C.Garen,
Y.Kolodenko,
A.E.Gorbalenya,
E.J.Snijder,
and
M.N.James
(2002).
Structure of arterivirus nsp4. The smallest chymotrypsin-like proteinase with an alpha/beta C-terminal extension and alternate conformations of the oxyanion hole.
|
| |
J Biol Chem,
277,
39960-39966.
|
|
|
PDB code:
|
|
|
|
|
|
|
The most recent references are shown first.
Citation data come partly from CiteXplore and partly
from an automated harvesting procedure. Note that this is likely to be
only a partial list as not all journals are covered by
either method. However, we are continually building up the citation data
so more and more references will be included with time.
Where a reference describes a PDB structure, the PDB
codes are
shown on the right.
|
|


 Links
Links































