Tryptophan synthase
Tryptophan synthase is able to catalyse the last two steps in the synthesis of tryptophan in bacteria, plants and fungi (but not in animals, where it is an essential component of the diet). It consists of two catalytically active subunits and was the first enzyme for which a product formed at one site was demonstrated to be intramolecularly transferred to another site.
The alpha subunit catalyses the cleavage of indolyl-glycerol-3-phosphate to give indole and 3-phosphoglyceraldehyde. Indole is then transferred to the beta subunit where it combines with L-serine to form tryptophan. This reaction is PLP-dependent.
Inhibitors of the alpha subunit are easy to design, meaning this enzyme represents a useful target for herbicides.
Reference Protein and Structure
- Sequences
-
P00929
 (4.2.1.20)
(4.2.1.20)
P0A2K1 (4.2.1.20)
 (Sequence Homologues)
(PDB Homologues)
(Sequence Homologues)
(PDB Homologues)
- Biological species
-
Salmonella enterica subsp. enterica serovar Typhimurium str. LT2 (Bacteria)
- PDB
-
1a50
- CRYSTAL STRUCTURE OF WILD-TYPE TRYPTOPHAN SYNTHASE COMPLEXED WITH 5-FLUOROINDOLE PROPANOL PHOSPHATE
(2.3 Å)



- Catalytic CATH Domains
-
3.20.20.70
 3.40.50.1100
(see all for 1a50)
3.40.50.1100
(see all for 1a50)
- Cofactors
- Pyridoxal 5'-phosphate(2-) (1)
Enzyme Reaction (EC:4.2.1.20)



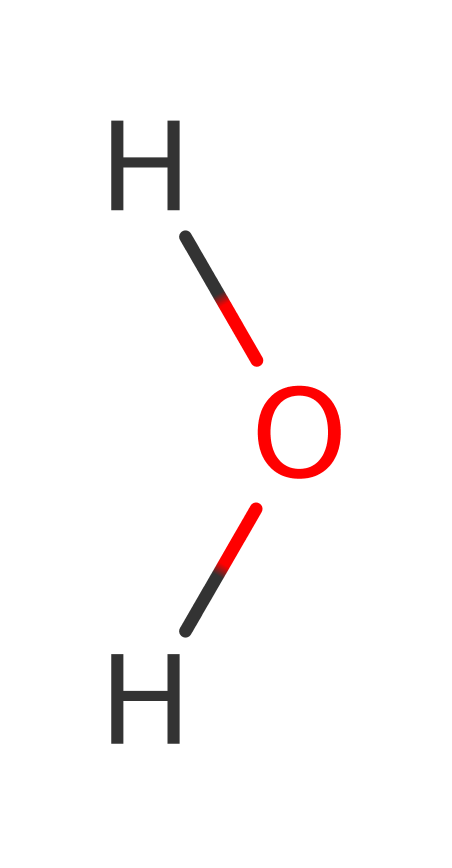

Enzyme Mechanism
- Summary
- Step 1
- Step 2
- Step 3
- Step 4
- Step 5
- Step 6
- Step 7
- Step 8
- Step 9
- Step 10
- Step 11
- Products
- All Steps
Introduction
The reaction takes place at two different sites, the α-site and β-site. The product from the alpha site is transferred through a tunnel to the beta site.
The α-active site catalyses the reversible formation of G3P and indole from IGP. The mechanism starts with protonation of the indole ring by Glu49 forming a charged reaction intermediate. The anion Glu49 abstracts a proton from the intermediate, triggering C-C bond cleavage to give indole and G3P. The indole molecule is transferred to the β-active site of the TSase.
In the β-active site, the catalytic activity requires a PLP cofactor which is covalently bound to an active site Lys. When the L-Ser substrate is present, a transimination reaction takes place in which it becomes covalently bound to PLP, forming an external aldimine (Aex-Ser). There are two parts to the reaction in the β-active site, a β-elimination and then a β-addition, which result in the tryptophan product covalently bonded to the PLP cofactor. From here, Aex-Trp undergoes a transimination reaction to form one molecule of tryptophan and Lys87 becomes covalently bonded to the PLP cofactor.
Catalytic Residues Roles
| UniProt | PDB* (1a50) | ||
| Glu49 | Glu49A | Protonated Glu49 interacts with the substrate via a water molecule. | hydrogen bond acceptor, proton acceptor, proton donor |
| Asp60 | Asp60A | Asp60 interacts closely with the substrate via two hydrogen bonds. | hydrogen bond acceptor |
| Lys87 | Lys87(86)B | Lys87 is involved in various proton transfers in the β-active site. | nucleofuge, nucleophile, proton acceptor, proton donor, electron pair acceptor, electron pair donor |
| Ser377 | Ser377(376)B | Ser377 stabilises the pyridine ring of PLP. | hydrogen bond donor |
| Thr183, Tyr102 | Thr183A, Tyr102A | Tyr102 and Thr183 stabilise Asp60 via hydrogen bonds. | hydrogen bond donor |
| Thr110 | Thr110(109)B | Thr110 stabilises the the carboxylate group of PLP. | hydrogen bond donor |
| Tyr175, Ser233, Ser235, Gly234 (main-N) | Tyr175A, Ser233A, Ser235A, Gly234A (main-N) | IGP is held by a network of hydrogen bonds formed by several residues in the α-active site. The interactions are centred on the phosphate group of IGP, anchoring the substrate in the α-active site. | hydrogen bond donor |
| Thr190, Ser235, Asn236, Gly232 (main-N), Gly233 (main-N), Gly234 (main-N) | Thr190(189)B, Ser235(234)B, Asn236(235)B, Gly232(231)B (main-N), Gly233(232)B (main-N), Gly234(233)B (main-N) | These residues provide a hydrogen bonding network throughout the β-active site mechanism. This mainly stabilises the phosphate group of Aex-Ser, fixing the position of PLP in the active site. | hydrogen bond donor |
Chemical Components
tautomerisation (not keto-enol), proton transfer, rate-determining step, heterolysis, native state of enzyme regenerated, intermediate formation, bimolecular nucleophilic addition, overall reactant used, inferred reaction step, unimolecular elimination by the conjugate base, cofactor used, atom stereo change, intramolecular elimination, bimolecular electrophilic addition, overall product formed, aromatic unimolecular elimination by the conjugate baseReferences
- Teixeira CSS et al. (2020), ChemCatChem, 12, 227-237. Solving the Catalytic Mechanism of Tryptophan Synthase: an Emergent Drug Target in the Treatment of Tuberculosis. DOI:10.1002/cctc.201901505.
- Huang YM et al. (2016), Protein Sci, 25, 166-183. Protonation states and catalysis: Molecular dynamics studies of intermediates in tryptophan synthase. DOI:10.1002/pro.2709. PMID:26013176.
- Caulkins BG et al. (2015), Biochim Biophys Acta, 1854, 1194-1199. Catalytic roles of βLys87 in tryptophan synthase: 15N solid state NMR studies. DOI:10.1016/j.bbapap.2015.02.003. PMID:25688830.
- Caulkins BG et al. (2014), J Am Chem Soc, 136, 12824-12827. Protonation States of the Tryptophan Synthase Internal Aldimine Active Site from Solid-State NMR Spectroscopy: Direct Observation of the Protonated Schiff Base Linkage to Pyridoxal-5′-Phosphate. DOI:10.1021/ja506267d. PMID:25148001.
- Lai J et al. (2011), J Am Chem Soc, 133, 4-7. X-ray and NMR Crystallography in an Enzyme Active Site: The Indoline Quinonoid Intermediate in Tryptophan Synthase. DOI:10.1021/ja106555c. PMID:21142052.
- Raboni S et al. (2009), Cell Mol Life Sci, 66, 2391-2403. Tryptophan synthase: a mine for enzymologists. DOI:10.1007/s00018-009-0028-0. PMID:19387555.
- Dunn MF et al. (2008), Trends Biochem Sci, 33, 254-264. Tryptophan synthase: the workings of a channeling nanomachine. DOI:10.1016/j.tibs.2008.04.008. PMID:18486479.
- Schiaretti F et al. (2004), J Biol Chem, 279, 29572-29582. pH Dependence of Tryptophan Synthase Catalytic Mechanism: I. THE FIRST STAGE, THE -ELIMINATION REACTION. DOI:10.1074/jbc.m401895200. PMID:15117965.
- Ferrari D et al. (2003), Biochemistry, 42, 7807-7818. Allosteric Communication in the Tryptophan Synthase Bienzyme Complex: Roles of the β-Subunit Aspartate 305−Arginine 141 Salt Bridge. DOI:10.1021/bi034291a. PMID:12820890.
- Miles EW (2001), Chem Rec, 1, 140-151. Tryptophan synthase: a multienzyme complex with an intramolecular tunnel. PMID:11893063.
- Ro HS et al. (1999), J Biol Chem, 274, 31189-31194. Catalytic Mechanism of the Tryptophan Synthase 2 2 Complex: EFFECTS OF pH, ISOTOPIC SUBSTITUTION, AND ALLOSTERIC LIGANDS. DOI:10.1074/jbc.274.44.31189. PMID:10531312.
- Sachpatzidis A et al. (1999), Biochemistry, 38, 12665-12674. Crystallographic Studies of Phosphonate-Based α-Reaction Transition-State Analogues Complexed to Tryptophan Synthase†,‡. DOI:10.1021/bi9907734. PMID:10504236.
- Weyand M et al. (1999), Biochemistry, 38, 16469-16480. Crystal structure of wild-type tryptophan synthase complexed with the natural substrate indole-3-glycerol phosphate. PMID:10600108.
- Schneider TR et al. (1998), Biochemistry, 37, 5394-5406. Loop Closure and Intersubunit Communication in Tryptophan Synthase†,‡. DOI:10.1021/bi9728957. PMID:9548921.
- Hyde CC et al. (1988), J Biol Chem, 263, 17857-17871. Three-dimensional structure of the tryptophan synthase alpha 2 beta 2 multienzyme complex from Salmonella typhimurium. PMID:3053720.

Step 1. In the α-active site there is a proton transfer from Glu49 to IGP via a water molecule. The substrate is held by a network of hydrogen bonds.
Download: Image, Marvin FileCatalytic Residues Roles
| Residue | Roles |
|---|---|
| Tyr175A | hydrogen bond donor |
| Thr183A | hydrogen bond donor |
| Tyr102A | hydrogen bond donor |
| Tyr173A | hydrogen bond donor |
| Glu49A | hydrogen bond acceptor |
| Asp60A | hydrogen bond acceptor |
| Glu49A | proton donor |
| Ser233A | hydrogen bond donor |
| Gly234A (main-N) | hydrogen bond donor |
| Ser235A | hydrogen bond donor |
Chemical Components
tautomerisation (not keto-enol), proton transfer, rate-determining step
Step 2. The anionic Glu49 abstracts a proton from the intermediate via a water molecule. This triggers C-C bond cleavage to give indole and G3P. The indole molecule is transferred to the β-active site of the enzyme via a tunnel. G3P remains in the α-subunit until the enzyme conformation changes to an open state.
Download: Image, Marvin FileCatalytic Residues Roles
| Residue | Roles |
|---|---|
| Glu49A | proton acceptor |
| Tyr175A | hydrogen bond donor |
| Ser233A | hydrogen bond donor |
| Gly234A (main-N) | hydrogen bond donor |
| Ser235A | hydrogen bond donor |
| Thr183A | hydrogen bond donor |
| Tyr102A | hydrogen bond donor |
| Tyr173A | hydrogen bond donor |
| Glu49A | hydrogen bond acceptor |
| Asp60A | hydrogen bond acceptor |
Chemical Components
proton transfer, heterolysis, native state of enzyme regenerated, intermediate formation
Step 3. In the β-active site, in the presence of the L-Ser substrate, a transimination reaction takes place in which the PLP cofactor is covalently bound to the substrate, forming an external aldimine (Aex-Ser). This step has not been studied in this enzyme but is shown here in a similar fashion to other PLP transamination reactions (see M-CSA ID: 66, for example).
Download: Image, Marvin FileCatalytic Residues Roles
| Residue | Roles |
|---|---|
| Lys87(86)B | proton acceptor, electron pair acceptor |
| Ser377(376)B | hydrogen bond donor |
| Thr190(189)B | hydrogen bond donor |
| Gly232(231)B (main-N) | hydrogen bond donor |
| Gly233(232)B (main-N) | hydrogen bond donor |
| Gly234(233)B (main-N) | hydrogen bond donor |
| Ser235(234)B | hydrogen bond donor |
| Asn236(235)B | hydrogen bond donor |
| Ser351(350)B | hydrogen bond donor |
| Thr110(109)B | hydrogen bond donor |
Chemical Components
ingold: bimolecular nucleophilic addition, proton transfer, overall reactant used, inferred reaction step
Step 4. Lys87 is eliminated from the covalent intermediate, leading to the formation of an external aldimine (Aex-Ser).
Download: Image, Marvin FileCatalytic Residues Roles
| Residue | Roles |
|---|---|
| Lys87(86)B | nucleofuge |
| Thr190(189)B | hydrogen bond donor |
| Gly232(231)B (main-N) | hydrogen bond donor |
| Gly233(232)B (main-N) | hydrogen bond donor |
| Gly234(233)B (main-N) | hydrogen bond donor |
| Ser235(234)B | hydrogen bond donor |
| Asn236(235)B | hydrogen bond donor |
| Ser351(350)B | hydrogen bond donor |
| Thr110(109)B | hydrogen bond donor |
| Ser377(376)B | hydrogen bond donor |
Chemical Components
ingold: unimolecular elimination by the conjugate base
Step 5. Abstraction of a proton from Aex-Ser by Lys87 forms a quinonoid intermediate.
Download: Image, Marvin FileCatalytic Residues Roles
| Residue | Roles |
|---|---|
| Thr190(189)B | hydrogen bond donor |
| Gly232(231)B (main-N) | hydrogen bond donor |
| Gly233(232)B (main-N) | hydrogen bond donor |
| Gly234(233)B (main-N) | hydrogen bond donor |
| Ser235(234)B | hydrogen bond donor |
| Asn236(235)B | hydrogen bond donor |
| Ser377(376)B | hydrogen bond donor |
| Thr110(109)B | hydrogen bond donor |
| Ser351(350)B | hydrogen bond donor |
Chemical Components
cofactor used, proton transfer, intermediate formation, rate-determining step
Step 6. The anionic intermediate is stabilised by the conjugated pi system provided by the PLP cofactor. Following dihedral rotation to align the hydroxyl group with Lys87, there is a proton transfer from Lys87 to the intermediate. Another intermediate is formed and a water molecule is released. Overall a C-C double bond forms.
Download: Image, Marvin FileCatalytic Residues Roles
| Residue | Roles |
|---|---|
| Ser377(376)B | hydrogen bond donor |
| Thr190(189)B | hydrogen bond donor |
| Gly232(231)B (main-N) | hydrogen bond donor |
| Gly233(232)B (main-N) | hydrogen bond donor |
| Gly234(233)B (main-N) | hydrogen bond donor |
| Ser235(234)B | hydrogen bond donor |
| Asn236(235)B | hydrogen bond donor |
| Thr110(109)B | hydrogen bond donor |
| Ser351(350)B | hydrogen bond donor |
Chemical Components
proton transfer, cofactor used, intermediate formation, atom stereo change, ingold: intramolecular elimination
Step 7. The indole molecule reaches the β-active site and is well aligned to react with the intermediate, forming a Trp quinonoid intermediate.
Download: Image, Marvin FileCatalytic Residues Roles
| Residue | Roles |
|---|---|
| Ser377(376)B | hydrogen bond donor |
| Thr190(189)B | hydrogen bond donor |
| Gly232(231)B (main-N) | hydrogen bond donor |
| Gly233(232)B (main-N) | hydrogen bond donor |
| Gly234(233)B (main-N) | hydrogen bond donor |
| Ser235(234)B | hydrogen bond donor |
| Asn236(235)B | hydrogen bond donor |
| Thr110(109)B | hydrogen bond donor |
Chemical Components
intermediate formation, ingold: bimolecular electrophilic addition
Step 8. Proton transfer to Lys87, forming a third Trp quinonoid reaction intermediate.
Download: Image, Marvin FileCatalytic Residues Roles
| Residue | Roles |
|---|---|
| Ser377(376)B | hydrogen bond donor |
| Thr190(189)B | hydrogen bond donor |
| Gly232(231)B (main-N) | hydrogen bond donor |
| Gly233(232)B (main-N) | hydrogen bond donor |
| Gly234(233)B (main-N) | hydrogen bond donor |
| Ser235(234)B | hydrogen bond donor |
| Asn236(235)B | hydrogen bond donor |
| Ser351(350)B | hydrogen bond donor |
| Thr110(109)B | hydrogen bond donor |
| Lys87(86)B | proton acceptor |
Chemical Components
rate-determining step, intermediate formation, proton transfer
Step 9. Proton transfer from Lys87 to the intermediate forming the tryptophan product covalently bonded to the PLP cofactor.
Download: Image, Marvin FileCatalytic Residues Roles
| Residue | Roles |
|---|---|
| Ser377(376)B | hydrogen bond donor |
| Thr190(189)B | hydrogen bond donor |
| Gly232(231)B (main-N) | hydrogen bond donor |
| Gly233(232)B (main-N) | hydrogen bond donor |
| Gly234(233)B (main-N) | hydrogen bond donor |
| Ser235(234)B | hydrogen bond donor |
| Asn236(235)B | hydrogen bond donor |
| Ser351(350)B | hydrogen bond donor |
| Thr110(109)B | hydrogen bond donor |
Chemical Components
proton transfer, overall product formed
Step 10. Aex-Trp undergoes a transamination reaction starting with a nucleophilic attack of Lys87 on the PLP cofactor.
Download: Image, Marvin FileCatalytic Residues Roles
| Residue | Roles |
|---|---|
| Lys87(86)B | nucleophile, proton donor |
| Ser377(376)B | hydrogen bond donor |
| Thr190(189)B | hydrogen bond donor |
| Gly232(231)B (main-N) | hydrogen bond donor |
| Gly233(232)B (main-N) | hydrogen bond donor |
| Gly234(233)B (main-N) | hydrogen bond donor |
| Ser235(234)B | hydrogen bond donor |
| Asn236(235)B | hydrogen bond donor |
| Ser351(350)B | hydrogen bond donor |
| Thr110(109)B | hydrogen bond donor |
Chemical Components
proton transfer, ingold: bimolecular nucleophilic addition
Step 11. The transamination reaction is completed to restore the Lys87 double bond to the PLP cofactor and produce a free tryptophan residue.
Download: Image, Marvin FileCatalytic Residues Roles
| Residue | Roles |
|---|---|
| Lys87(86)B | electron pair donor |
| Ser377(376)B | hydrogen bond donor |
| Thr190(189)B | hydrogen bond donor |
| Gly232(231)B (main-N) | hydrogen bond donor |
| Gly233(232)B (main-N) | hydrogen bond donor |
| Gly234(233)B (main-N) | hydrogen bond donor |
| Ser235(234)B | hydrogen bond donor |
| Asn236(235)B | hydrogen bond donor |
| Ser351(350)B | hydrogen bond donor |
| Thr110(109)B | hydrogen bond donor |
 Download:
Download: