 |
PDBsum entry 2nya

|
|
|
|
|
|

|

|

|

|
|

|
|

|
|

|
|

|
|

|
|

|
|

|
|
|
 |

|

|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Oxidoreductase
|
PDB id
|
|
|
|
2nya
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|

|

|

|
|
|
|
|
|
|
|
 |
Contents |
 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
* Residue conservation analysis
|
|
|
|
|
|
|
|
|
|
|
Enzyme class:
|
|
E.C.1.9.6.1
- nitrate reductase (cytochrome).
|
|
|
|
|
|
|
Reaction:
|
|
2 Fe(II)-[cytochrome] + nitrate + 2 H+ = 2 Fe(III)-[cytochrome] + nitrite + H2O
|
|
|
|
|
|
2
×
Fe(II)-[cytochrome]
|
+
|
 nitrate
nitrate
|
+
|
2
×
H(+)
|
=
|
2
×
Fe(III)-[cytochrome]
|
+
|
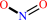 nitrite
nitrite
|
+
|
H2O
|
|
|
|
|
|
|
|
|
|
|
|
|
Molecule diagrams generated from .mol files obtained from the
KEGG ftp site
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
| |
|
|
| |
|
DOI no:
|
J Biol Chem
282:6425-6437
(2007)
|
|
PubMed id:
|
|
|
|
|
|
| |
|
Spectropotentiometric and structural analysis of the periplasmic nitrate reductase from Escherichia coli.
|
|
B.J.Jepson,
S.Mohan,
T.A.Clarke,
A.J.Gates,
J.A.Cole,
C.S.Butler,
J.N.Butt,
A.M.Hemmings,
D.J.Richardson.
|
|
|
|
|
| |
ABSTRACT
|
|
|
|
| |
|
|
The Escherichia coli NapA (periplasmic nitrate reductase) contains a [4Fe-4S]
cluster and a Mo-bis-molybdopterin guanine dinucleotide cofactor. The NapA
holoenzyme associates with a di-heme c-type cytochrome redox partner (NapB).
These proteins have been purified and studied by spectropotentiometry, and the
structure of NapA has been determined. In contrast to the well characterized
heterodimeric NapAB systems ofalpha-proteobacteria, such as Rhodobacter
sphaeroides and Paracoccus pantotrophus, the gamma-proteobacterial E. coli NapA
and NapB proteins purify independently and not as a tight heterodimeric complex.
This relatively weak interaction is reflected in dissociation constants of 15
and 32 mum determined for oxidized and reduced NapAB complexes, respectively.
The surface electrostatic potential of E. coli NapA in the apparent NapB binding
region is markedly less polar and anionic than that of the alpha-proteobacterial
NapA, which may underlie the weaker binding of NapB. The molybdenum ion
coordination sphere of E. coli NapA includes two molybdopterin guanine
dinucleotide dithiolenes, a protein-derived cysteinyl ligand and an oxygen atom.
The Mo-O bond length is 2.6 A, which is indicative of a water ligand. The
potential range over which the Mo(6+) state is reduced to the Mo(5+) state in
either NapA (between +100 and -100 mV) or the NapAB complex (-150 to -350 mV) is
much lower than that reported for R. sphaeroides NapA (midpoint potential
Mo(6+/5+) > +350 mV), and the form of the Mo(5+) EPR signal is quite distinct.
In E. coli NapA or NapAB, the Mo(5+) state could not be further reduced to
Mo(4+). We then propose a catalytic cycle for E. coli NapA in which nitrate
binds to the Mo(5+) ion and where a stable des-oxo Mo(6+) species may
participate.
|
|
|
|
|
|
| |
Selected figure(s)
|
|
|
|
| |
|
|

|
|

|
|
Figure 5.
FIGURE 5. Crystal structure of E. coli NapA. A, view of
NapA from E. coli with the different domains colored as follows:
domain I (residues 1–59, 492–520, and 590–630) in red;
domain II (residues 60–138, 375–491, and 521–589) in
green; domain III (residues 139–374) in yellow; and domain IV
(residues 631–791) in blue. The regions colored gray are the
extra loop regions not present in the D. desulfuricans NAP (9).
The [4Fe-4S] cluster and the Mo-bis-MGD cofactors are shown in
ball and stick format and are colored black. B, view of the
4Fe4S cluster and Mo-bis-MGD cofactor showing the conserved
water molecule that is coordinated by Lys-47 and one of the MGD
moieties.
|
|
Figure 6.
FIGURE 6. Electrostatic surface representations of the NAP
structures. A, electrostatic surface of the molybdenum funnel
face of the various NAP structures. B, electrostatic surface of
the presumed NapB interaction face of the various NAP
structures. Electrostatic surfaces were calculated in the
program GRASP (28) and displayed in PYMOL (Delano Scientific).
The "surface potentials" of color property scale -10 (red,
negatively charged) to +10 (blue, positively charged).
Coordinates for D. desulfuricans NAP, PDB file 2NAP. Coordinates
for R. sphaeroides NapA, Protein Data Bank (PDB) file 1OGY. The
structures of E. coli, D. desulfuricans, and R. sphaeroides were
aligned using Swisspdbviewer 3.0 ("Magic Fit"). D.d, D.
desulfuricans; E.c, E. coli, R.s, R. sphaeroides.
|
|
|
|
|
|
| |
The above figures are
reprinted
by permission from the ASBMB:
J Biol Chem
(2007,
282,
6425-6437)
copyright 2007.
|
|
| |
Figures were
selected
by the author.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
 |
 |
|
Literature references that cite this PDB file's key reference
|
|
|
| |
PubMed id
|
|
Reference
|
|
|
|
|
|
A.J.Gates,
G.L.Kemp,
C.Y.To,
J.Mann,
S.J.Marritt,
A.G.Mayes,
D.J.Richardson,
and
J.N.Butt
(2011).
The relationship between redox enzyme activity and electrochemical potential-cellular and mechanistic implications from protein film electrochemistry.
|
| |
Phys Chem Chem Phys,
13,
7720-7731.
|
|
|
|
|
|
|
A.J.Gates,
V.M.Luque-Almagro,
A.D.Goddard,
S.J.Ferguson,
M.D.Roldán,
and
D.J.Richardson
(2011).
A composite biochemical system for bacterial nitrate and nitrite assimilation as exemplified by Paracoccus denitrificans.
|
| |
Biochem J,
435,
743-753.
|
|
|
|
|
|
|
C.Coelho,
P.J.González,
J.G.Moura,
I.Moura,
J.Trincão,
and
M.João Romão
(2011).
The crystal structure of Cupriavidus necator nitrate reductase in oxidized and partially reduced states.
|
| |
J Mol Biol,
408,
932-948.
|
|
|
PDB codes:
|
|
|
|
|
|
|
|
A.A.Filimonenkov,
R.A.Zvyagilskaya,
T.V.Tikhonova,
and
V.O.Popov
(2010).
Isolation and characterization of nitrate reductase from the halophilic sulfur-oxidizing bacterium Thioalkalivibrio nitratireducens.
|
| |
Biochemistry (Mosc),
75,
744-751.
|
|
|
|
|
|
|
P.J.Simpson,
D.J.Richardson,
and
R.Codd
(2010).
The periplasmic nitrate reductase in Shewanella: the resolution, distribution and functional implications of two NAP isoforms, NapEDABC and NapDAGHB.
|
| |
Microbiology,
156,
302-312.
|
|
|
|
|
|
|
H.Gao,
Z.K.Yang,
S.Barua,
S.B.Reed,
M.F.Romine,
K.H.Nealson,
J.K.Fredrickson,
J.M.Tiedje,
and
J.Zhou
(2009).
Reduction of nitrate in Shewanella oneidensis depends on atypical NAP and NRF systems with NapB as a preferred electron transport protein from CymA to NapA.
|
| |
ISME J,
3,
966-976.
|
|
|
|
|
|
|
M.Hofmann
(2009).
Density functional theory study of model complexes for the revised nitrate reductase active site in Desulfovibrio desulfuricans NapA.
|
| |
J Biol Inorg Chem,
14,
1023-1035.
|
|
|
|
|
|
|
M.J.Romão
(2009).
Molybdenum and tungsten enzymes: a crystallographic and mechanistic overview.
|
| |
Dalton Trans,
(),
4053-4068.
|
|
|
|
|
|
|
N.M.Cerqueira,
P.J.Gonzalez,
C.D.Brondino,
M.J.Romão,
C.C.Romão,
I.Moura,
and
J.J.Moura
(2009).
The effect of the sixth sulfur ligand in the catalytic mechanism of periplasmic nitrate reductase.
|
| |
J Comput Chem,
30,
2466-2484.
|
|
|
|
|
|
|
S.Najmudin,
P.J.González,
J.Trincão,
C.Coelho,
A.Mukhopadhyay,
N.M.Cerqueira,
C.C.Romão,
I.Moura,
J.J.Moura,
C.D.Brondino,
and
M.J.Romão
(2008).
Periplasmic nitrate reductase revisited: a sulfur atom completes the sixth coordination of the catalytic molybdenum.
|
| |
J Biol Inorg Chem,
13,
737-753.
|
|
|
PDB codes:
|
|
|
|
|
|
|
|
C.Coelho,
P.J.González,
J.Trincão,
A.L.Carvalho,
S.Najmudin,
T.Hettman,
S.Dieckman,
J.J.Moura,
I.Moura,
and
M.J.Romão
(2007).
Heterodimeric nitrate reductase (NapAB) from Cupriavidus necator H16: purification, crystallization and preliminary X-ray analysis.
|
| |
Acta Crystallogr Sect F Struct Biol Cryst Commun,
63,
516-519.
|
|
|
|
|
|
|
J.Maillard,
C.A.Spronk,
G.Buchanan,
V.Lyall,
D.J.Richardson,
T.Palmer,
G.W.Vuister,
and
F.Sargent
(2007).
Structural diversity in twin-arginine signal peptide-binding proteins.
|
| |
Proc Natl Acad Sci U S A,
104,
15641-15646.
|
|
|
PDB code:
|
|
|
|
|
|
|
The most recent references are shown first.
Citation data come partly from CiteXplore and partly
from an automated harvesting procedure. Note that this is likely to be
only a partial list as not all journals are covered by
either method. However, we are continually building up the citation data
so more and more references will be included with time.
Where a reference describes a PDB structure, the PDB
codes are
shown on the right.
|
|



 Links
Links














