 |
PDBsum entry 1a5u

|
|
|
|
|
 |
Contents |
 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
* Residue conservation analysis
|
|
|
|
|
|
PDB id:
|
|
|
|
| Name: |
|
Transferase
|
|
|
Title:
|
|
Pyruvate kinase complex with bis mg-atp-na-oxalate
|
|
Structure:
|
|
Pyruvate kinase. Chain: a, b, c, d, e, f, g, h. Ec: 2.7.1.40
|
|
Source:
|
|
Oryctolagus cuniculus. Rabbit. Organism_taxid: 9986. Tissue: muscle
|
|
Biol. unit:
|
|
 Tetramer (from PDB file)
Tetramer (from PDB file)
|
|
Resolution:
|
|
|
|
Authors:
|
|
T.M.Larsen,M.M.Benning,I.Rayment,G.H.Reed
|
Key ref:

|
|
T.M.Larsen
et al.
(1998).
Structure of the bis(Mg2+)-ATP-oxalate complex of the rabbit muscle pyruvate kinase at 2.1 A resolution: ATP binding over a barrel.
Biochemistry,
37,
6247-6255.
PubMed id:
DOI:
|
|
|
Date:
|
|
|
18-Feb-98
|
Release date:
|
02-Mar-99
|
|
|
|
|
|
|
PROCHECK
|

|
|
|
|
|
Headers
|
|
|
|
References
|
|
|
|
|
|

|
|
P11974
(KPYM_RABIT) -
Pyruvate kinase PKM from Oryctolagus cuniculus
|
|
|
|
Seq:
Struc:
|
|
|
|
531 a.a.
519 a.a.
|
|

|

|
|
|
|
|
|
|
|
|
|
|
|
Key: |
 |
PfamA domain |
|
|
 |
Secondary structure |
|
 |
CATH domain |
|
|
|
|
|
|
|
|
|
|
|
|
|
Enzyme class 2:
|
|
E.C.2.7.1.40
- pyruvate kinase.
|
|
|
|
|
|
|
Reaction:
|
|
pyruvate + ATP = phosphoenolpyruvate + ADP + H+
|
|
|
|
|
|
pyruvate
Bound ligand (Het Group name = )
corresponds exactly
|
+
|
ATP
Bound ligand (Het Group name = )
matches with 71.43% similarity
|
=
|
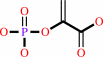 phosphoenolpyruvate
phosphoenolpyruvate
|
+
|
 ADP
ADP
|
+
|
H(+)
|
|
|
|
|
|
|
|
|
|
Enzyme class 3:
|
|
E.C.2.7.10.2
- non-specific protein-tyrosine kinase.
|
|
|
|
|
|
|
Reaction:
|
|
L-tyrosyl-[protein] + ATP = O-phospho-L-tyrosyl-[protein] + ADP + H+
|
|
|
|
|
|
L-tyrosyl-[protein]
Bound ligand (Het Group name = )
corresponds exactly
|
+
|
 ATP
ATP
|
=
|
O-phospho-L-tyrosyl-[protein]
|
+
|
ADP
|
+
|
H(+)
|
|
|
|
|
|
|
|
|
|
Enzyme class 4:
|
|
E.C.2.7.11.1
- non-specific serine/threonine protein kinase.
|
|
|
|
|
|
|
Reaction:
|
|
|
1.
|
L-seryl-[protein] + ATP = O-phospho-L-seryl-[protein] + ADP + H+
|
|
2.
|
L-threonyl-[protein] + ATP = O-phospho-L-threonyl-[protein] + ADP + H+
|
|
|
|
|
|
|
L-seryl-[protein]
Bound ligand (Het Group name = )
corresponds exactly
|
+
|
ATP
|
=
|
O-phospho-L-seryl-[protein]
|
+
|
ADP
|
+
|
H(+)
|
|
|
|
|
|
|
L-threonyl-[protein]
Bound ligand (Het Group name = )
corresponds exactly
|
+
|
ATP
|
=
|
O-phospho-L-threonyl-[protein]
|
+
|
ADP
|
+
|
H(+)
|
|
|
|
|
|
|
|
|
|
|
|
|
Note, where more than one E.C. class is given (as above), each may
correspond to a different protein domain or, in the case of polyprotein
precursors, to a different mature protein.
|
|
|
|
Molecule diagrams generated from .mol files obtained from the
KEGG ftp site
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
| |
|
|
| |
|
DOI no:
|
Biochemistry
37:6247-6255
(1998)
|
|
PubMed id:
|
|
|
|
|
|
| |
|
Structure of the bis(Mg2+)-ATP-oxalate complex of the rabbit muscle pyruvate kinase at 2.1 A resolution: ATP binding over a barrel.
|
|
T.M.Larsen,
M.M.Benning,
I.Rayment,
G.H.Reed.
|
|
|
|
|
| |
ABSTRACT
|
|
|
|
| |
|
|
Pyruvate kinase from rabbit muscle has been cocrystallized as a complex with
MgIIATP, oxalate, Mg2+, and either K+ or Na+. Crystals with either Na+ or K+
belong to the space group P2(1)2(1)2(1), and the asymmetric units contain two
tetramers. The structures were solved by molecular replacement and refined to
2.1 (K+) and 2.35 A (Na+) resolution. The structures of the Na+ and K+ complexes
are virtually isomorphous. Each of the eight subunits within the asymmetric unit
contains MgIIoxalate as a bidentate complex linked to the protein through
coordination of Mg2+ to the carboxylates of Glu 271 and Asp 295. Six of the
subunits also contain an alpha,beta,gamma-tridentate complex of MgIIATP, and the
active-site cleft, located between domains A and B, is closed in these subunits.
In the remaining two subunits MgIIATP is missing, and the active-site cleft is
open. Closure of the active-site cleft in the fully liganded subunits includes a
rotation of 41 degrees of the B domain relative to the A domain. alpha-Carbons
of residues in the B domain undergo movements of up to 17.8 A (Lys 124) in the
cleft closure. Lys 206, Arg 119, and Asp 177 from the B domain move several
angstroms from their positions in the open conformation to contact the MgIIATP
complex in the active site. The gamma-phosphate of ATP coordinates to both
magnesium ions and to the monovalent cation, K+ or Na+. A Mg2+-coordinated
oxygen from the MgIIoxalate complex lies 3.0 A from Pgamma of ATP, and this
oxygen is positioned for an in-line attack on the phosphorus. The side chains of
Lys 269 and Arg 119 are positioned to provide leaving-group activation in the
forward and reverse directions. There is no obvious candidate for the acid/base
catalyst near the 2-si face of the prospective enolate of the normal substrate.
A functional group linked through solvent and side-chain hydroxyls may function
in a proton relay.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
 |
 |
|
Literature references that cite this PDB file's key reference
|
|
|
| |
PubMed id
|
|
Reference
|
|
|
|
|
|
A.Kahraman,
R.J.Morris,
R.A.Laskowski,
A.D.Favia,
and
J.M.Thornton
(2010).
On the diversity of physicochemical environments experienced by identical ligands in binding pockets of unrelated proteins.
|
| |
Proteins,
78,
1120-1136.
|
|
|
|
|
|
|
M.J.Lee,
L.Chien-Liang,
J.Y.Tsai,
W.T.Sue,
W.S.Hsia,
and
H.Huang
(2010).
Identification and biochemical characterization of a unique Mn2+-dependent UMP kinase from Helicobacter pylori.
|
| |
Arch Microbiol,
192,
739-746.
|
|
|
|
|
|
|
R.Bakszt,
A.Wernimont,
A.Allali-Hassani,
M.W.Mok,
T.Hills,
R.Hui,
and
J.C.Pizarro
(2010).
The crystal structure of Toxoplasma gondii pyruvate kinase 1.
|
| |
PLoS One,
5,
e12736.
|
|
|
PDB codes:
|
|
|
|
|
|
|
|
S.Kumar,
and
A.Barth
(2010).
Phosphoenolpyruvate and Mg2+ binding to pyruvate kinase monitored by infrared spectroscopy.
|
| |
Biophys J,
98,
1931-1940.
|
|
|
|
|
|
|
G.D.Markham,
F.Takusagawa,
A.M.Dijulio,
and
C.W.Bock
(2009).
An investigation of the catalytic mechanism of S-adenosylmethionine synthetase by QM/MM calculations.
|
| |
Arch Biochem Biophys,
492,
82-92.
|
|
|
|
|
|
|
R.van Wijk,
E.G.Huizinga,
A.C.van Wesel,
B.A.van Oirschot,
M.A.Hadders,
and
W.W.van Solinge
(2009).
Fifteen novel mutations in PKLR associated with pyruvate kinase (PK) deficiency: structural implications of amino acid substitutions in PK.
|
| |
Hum Mutat,
30,
446-453.
|
|
|
|
|
|
|
J.C.Lee
(2008).
Modulation of allostery of pyruvate kinase by shifting of an ensemble of microstates.
|
| |
Acta Biochim Biophys Sin (Shanghai),
40,
663-669.
|
|
|
|
|
|
|
J.Petersen,
K.Fisher,
and
D.J.Lowe
(2008).
Structural basis for VO2+ inhibition of nitrogenase activity (A): 31P and 23Na interactions with the metal at the nucleotide binding site of the nitrogenase Fe protein identified by ENDOR spectroscopy.
|
| |
J Biol Inorg Chem,
13,
623-635.
|
|
|
|
|
|
|
S.Varma,
and
S.B.Rempe
(2008).
Structural transitions in ion coordination driven by changes in competition for ligand binding.
|
| |
J Am Chem Soc,
130,
15405-15419.
|
|
|
|
|
|
|
F.N.Musayev,
M.L.di Salvo,
T.P.Ko,
A.K.Gandhi,
A.Goswami,
V.Schirch,
and
M.K.Safo
(2007).
Crystal Structure of human pyridoxal kinase: structural basis of M(+) and M(2+) activation.
|
| |
Protein Sci,
16,
2184-2194.
|
|
|
PDB codes:
|
|
|
|
|
|
|
|
A.Müller,
L.Toma,
H.Bögge,
M.Henry,
E.T.Haupt,
A.Mix,
and
F.L.Sousa
(2006).
Reactions inside a porous nanocapsule/artificial cell: encapsulates' structuring directed by internal surface deprotonations.
|
| |
Chem Commun (Camb),
(),
3396-3398.
|
|
|
|
|
|
|
E.Di Cera
(2006).
A structural perspective on enzymes activated by monovalent cations.
|
| |
J Biol Chem,
281,
1305-1308.
|
|
|
|
|
|
|
J.Oria-Hernández,
H.Riveros-Rosas,
and
L.Ramírez-Sílva
(2006).
Dichotomic phylogenetic tree of the pyruvate kinase family: K+ -dependent and -independent enzymes.
|
| |
J Biol Chem,
281,
30717-30724.
|
|
|
|
|
|
|
F.Glaser,
Y.Rosenberg,
A.Kessel,
T.Pupko,
and
N.Ben-Tal
(2005).
The ConSurf-HSSP database: the mapping of evolutionary conservation among homologs onto PDB structures.
|
| |
Proteins,
58,
610-617.
|
|
|
|
|
|
|
J.Oria-Hernández,
N.Cabrera,
R.Pérez-Montfort,
and
L.Ramírez-Silva
(2005).
Pyruvate kinase revisited: the activating effect of K+.
|
| |
J Biol Chem,
280,
37924-37929.
|
|
|
|
|
|
|
N.Fernandez-Fuentes,
A.Hermoso,
J.Espadaler,
E.Querol,
F.X.Aviles,
and
B.Oliva
(2004).
Classification of common functional loops of kinase super-families.
|
| |
Proteins,
56,
539-555.
|
|
|
|
|
|
|
B.N.Chaudhuri,
M.R.Sawaya,
C.Y.Kim,
G.S.Waldo,
M.S.Park,
T.C.Terwilliger,
and
T.O.Yeates
(2003).
The crystal structure of the first enzyme in the pantothenate biosynthetic pathway, ketopantoate hydroxymethyltransferase, from M tuberculosis.
|
| |
Structure,
11,
753-764.
|
|
|
PDB code:
|
|
|
|
|
|
|
|
D.Susan-Resiga,
and
T.Nowak
(2003).
The proton transfer step catalyzed by yeast pyruvate kinase.
|
| |
J Biol Chem,
278,
12660-12671.
|
|
|
|
|
|
|
F.Schmitzberger,
A.G.Smith,
C.Abell,
and
T.L.Blundell
(2003).
Comparative analysis of the Escherichia coli ketopantoate hydroxymethyltransferase crystal structure confirms that it is a member of the (betaalpha)8 phosphoenolpyruvate/pyruvate superfamily.
|
| |
J Bacteriol,
185,
4163-4171.
|
|
|
|
|
|
|
L.Ramírez-Silva,
and
J.Oria-Hernández
(2003).
Selectivity of pyruvate kinase for Na+ and K+ in water/dimethylsulfoxide mixtures.
|
| |
Eur J Biochem,
270,
2377-2385.
|
|
|
|
|
|
|
G.Valentini,
L.R.Chiarelli,
R.Fortin,
M.Dolzan,
A.Galizzi,
D.J.Abraham,
C.Wang,
P.Bianchi,
A.Zanella,
and
A.Mattevi
(2002).
Structure and function of human erythrocyte pyruvate kinase. Molecular basis of nonspherocytic hemolytic anemia.
|
| |
J Biol Chem,
277,
23807-23814.
|
|
|
PDB codes:
|
|
|
|
|
|
|
|
L.F.Garcia-Alles,
K.Flükiger,
J.Hewel,
R.Gutknecht,
C.Siebold,
S.Schürch,
and
B.Erni
(2002).
Mechanism-based inhibition of enzyme I of the Escherichia coli phosphotransferase system. Cysteine 502 is an essential residue.
|
| |
J Biol Chem,
277,
6934-6942.
|
|
|
|
|
|
|
S.Ramón-Maiques,
A.Marina,
F.Gil-Ortiz,
I.Fita,
and
V.Rubio
(2002).
Structure of acetylglutamate kinase, a key enzyme for arginine biosynthesis and a prototype for the amino acid kinase enzyme family, during catalysis.
|
| |
Structure,
10,
329-342.
|
|
|
PDB codes:
|
|
|
|
|
|
|
|
A.Heine,
G.DeSantis,
J.G.Luz,
M.Mitchell,
C.H.Wong,
and
I.A.Wilson
(2001).
Observation of covalent intermediates in an enzyme mechanism at atomic resolution.
|
| |
Science,
294,
369-374.
|
|
|
PDB codes:
|
|
|
|
|
|
|
|
L.Ramírez-Silva,
S.T.Ferreira,
T.Nowak,
M.Tuena de Gómez-Puyou,
and
A.Gómez-Puyou
(2001).
Dimethylsulfoxide promotes K+-independent activity of pyruvate kinase and the acquisition of the active catalytic conformation.
|
| |
Eur J Biochem,
268,
3267-3274.
|
|
|
|
|
|
|
A.Schramm,
B.Siebers,
B.Tjaden,
H.Brinkmann,
and
R.Hensel
(2000).
Pyruvate kinase of the hyperthermophilic crenarchaeote Thermoproteus tenax: physiological role and phylogenetic aspects.
|
| |
J Bacteriol,
182,
2001-2009.
|
|
|
|
|
|
|
H.Erlandsen,
E.E.Abola,
and
R.C.Stevens
(2000).
Combining structural genomics and enzymology: completing the picture in metabolic pathways and enzyme active sites.
|
| |
Curr Opin Struct Biol,
10,
719-730.
|
|
|
|
|
|
|
T.Zhou,
S.Radaev,
B.P.Rosen,
and
D.L.Gatti
(2000).
Structure of the ArsA ATPase: the catalytic subunit of a heavy metal resistance pump.
|
| |
EMBO J,
19,
4838-4845.
|
|
|
PDB code:
|
|
|
|
|
|
|
|
B.A.Bernat,
L.T.Laughlin,
and
R.N.Armstrong
(1999).
Elucidation of a monovalent cation dependence and characterization of the divalent cation binding site of the fosfomycin resistance protein (FosA).
|
| |
Biochemistry,
38,
7462-7469.
|
|
|
|
|
|
|
T.J.Bollenbach,
A.D.Mesecar,
and
T.Nowak
(1999).
Role of lysine 240 in the mechanism of yeast pyruvate kinase catalysis.
|
| |
Biochemistry,
38,
9137-9145.
|
|
|
|
|
|
|
T.Zhou,
and
B.P.Rosen
(1999).
Asp45 is a Mg2+ ligand in the ArsA ATPase.
|
| |
J Biol Chem,
274,
13854-13858.
|
|
|
|
|
|
The most recent references are shown first.
Citation data come partly from CiteXplore and partly
from an automated harvesting procedure. Note that this is likely to be
only a partial list as not all journals are covered by
either method. However, we are continually building up the citation data
so more and more references will be included with time.
Where a reference describes a PDB structure, the PDB
codes are
shown on the right.
|
|




 Links
Links







