
|
|
|
|
|
 |
Contents |
 |
|
|
|
|
|
|
|
|
|
|
* Residue conservation analysis
|
|
|
|
|
|
|
|
|
|
Enzyme class 2:
|
|
Chain A:
E.C.2.7.1.197
- protein-N(pi)-phosphohistidine--D-mannitol phosphotransferase.
|
|
|
|
|
|
|
Reaction:
|
|
D-mannitol(out) + N(pros)-phospho-L-histidyl-[protein] = D-mannitol 1-phosphate(in) + L-histidyl-[protein]
|
|
|
|
|
|
[Protein]-N(pi)-phospho-L-histidine
|
+
|
D-mannitol(Side 1)
|
=
|
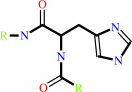 [protein]-L- histidine
[protein]-L- histidine
|
+
|
D-mannitol 1-phosphate(Side 2)
|
|
|
|
|
|
|
|
|
|
Enzyme class 3:
|
|
Chain B:
E.C.?
|
|
|
|
|
|
|
|
|
|
Note, where more than one E.C. class is given (as above), each may
correspond to a different protein domain or, in the case of polyprotein
precursors, to a different mature protein.
|
|
|
|
Molecule diagrams generated from .mol files obtained from the
KEGG ftp site
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
| |
|
|
| |
|
DOI no:
|
J Biol Chem
277:42289-42298
(2002)
|
|
PubMed id:
|
|
|
|
|
|
| |
|
Solution structure of the phosphoryl transfer complex between the cytoplasmic A domain of the mannitol transporter IIMannitol and HPr of the Escherichia coli phosphotransferase system.
|
|
G.Cornilescu,
B.R.Lee,
C.C.Cornilescu,
G.Wang,
A.Peterkofsky,
G.M.Clore.
|
|
|
|
|
| |
ABSTRACT
|
|
|
|
| |
|
|
The solution structure of the complex between the cytoplasmic A domain
(IIA(Mtl)) of the mannitol transporter II(Mannitol) and the histidine-containing
phosphocarrier protein (HPr) of the Escherichia coli phosphotransferase system
has been solved by NMR, including the use of conjoined rigid body/torsion angle
dynamics, and residual dipolar couplings, coupled with cross-validation, to
permit accurate orientation of the two proteins. A convex surface on HPr, formed
by helices 1 and 2, interacts with a complementary concave depression on the
surface of IIA(Mtl) formed by helix 3, portions of helices 2 and 4, and
beta-strands 2 and 3. The majority of intermolecular contacts are hydrophobic,
with a small number of electrostatic interactions at the periphery of the
interface. The active site histidines, His-15 of HPr and His-65 of IIA(Mtl), are
in close spatial proximity, and a pentacoordinate phosphoryl transition state
can be readily accommodated with no change in protein-protein orientation and
only minimal perturbations of the backbone immediately adjacent to the
histidines. Comparison with two previously solved structures of complexes of HPr
with partner proteins of the phosphotransferase system, the N-terminal domain of
enzyme I (EIN) and enzyme IIA(Glucose) (IIA(Glc)), reveals a number of common
features despite the fact that EIN, IIA(Glc), and IIA(Mtl) bear no structural
resemblance to one another. Thus, entirely different underlying structural
elements can form binding surfaces for HPr that are similar in terms of both
shape and residue composition. These structural comparisons illustrate the roles
of surface and residue complementarity, redundancy, incremental build-up of
specificity and conformational side chain plasticity in the formation of
transient specific protein-protein complexes in signal transduction pathways.
|
|
|
|
|
|
| |
Selected figure(s)
|
|
|
|
| |
|
|

|
|

|
|
Figure 5.
Fig. 5. The transition state of the IIA^Mtl-P-HPr
complex. A, detailed view around the active site histidines,
illustrating the backbone and side chain positions in the
unphosphorylated complex, the dissociative transition state, and
the associative transition state. The backbones of IIA^Mtl and
HPr are shown in dark blue and dark green, respectively, for the
unphosphorylated complex, and in light blue and light green,
respectively, for the putative dissociative and associative
transition states; the active site histidines and
pentacoordinate phosphoryl group (in the case of the transition
states) are shown in purple for the unphosphorylated complex, in
red for the dissociative transition state (N  2-N 2-N  1 distance
of ~6 Å between His-65 and His-15), and in orange for the
associative transition state (N 2-N 1 distance
of ~4 Å between His-65 and His-15). Small changes in the
backbone of residues 64-66 of IIA^Mtl and residues 14-16 of HPr
are required to accommodate the transition states. B, detailed
view of the active site in the putative transition state
illustrating the interactions that stabilize the phosphoryl
group. The color coding is as follows: the backbone and side
chains of IIA^Mtl are shown in blue and red, respectively; the
backbone and side chains of HPr are shown in green and gray,
respectively; the active site histidines are in purple, and the
pentacoordinate phosphoryl group is in yellow. Residues from HPr
are labeled in italics. 1 distance
of ~6 Å between His-65 and His-15), and in orange for the
associative transition state (N 2-N 1 distance
of ~4 Å between His-65 and His-15). Small changes in the
backbone of residues 64-66 of IIA^Mtl and residues 14-16 of HPr
are required to accommodate the transition states. B, detailed
view of the active site in the putative transition state
illustrating the interactions that stabilize the phosphoryl
group. The color coding is as follows: the backbone and side
chains of IIA^Mtl are shown in blue and red, respectively; the
backbone and side chains of HPr are shown in green and gray,
respectively; the active site histidines are in purple, and the
pentacoordinate phosphoryl group is in yellow. Residues from HPr
are labeled in italics.
|
|
Figure 6.
Fig. 6. Surface representations illustrating the binding
surfaces involved in the IIA^Mtl-HPr (left panel), IIA^Glc-HPr
(middle panel), and EIN-HPr (right panel) complexes. The binding
surfaces on IIA^Mtl, IIA Glc, and EIN are shown in A, and the
binding surfaces on HPr are shown in B. The binding surfaces are
color-coded with hydrophobic residues in green, polar residues
in light blue, the active site histidines in purple, positively
charged residues in dark blue, and negatively charged residues
in red. The relevant portions of the backbone of HPr are shown
in gold in A; the relevant portions of the backbone of IIA^Mtl,
IIA^Glc, and EIN are shown in blue, red, and green,
respectively, in B. The location of the phosphoryl group in the
transition state is depicted in yellow. Residues of HPr are
labeled in italics. The coordinates of the EIN-HPr and
IIA^Glc-HPr complexes are taken from Garrett et al. (16) (code
3EZE) and Wang et al. (17) (code 1GGR), respectively.
|
|
|
|
|
|
| |
The above figures are
reprinted
by permission from the ASBMB:
J Biol Chem
(2002,
277,
42289-42298)
copyright 2002.
|
|
| |
Figures were
selected
by an automated process.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
 |
 |
|
Literature references that cite this PDB file's key reference
|
|
|
| |
PubMed id
|
|
Reference
|
|
|
|
|
|
K.Pflüger-Grau,
and
B.Görke
(2010).
Regulatory roles of the bacterial nitrogen-related phosphotransferase system.
|
| |
Trends Microbiol,
18,
205-214.
|
|
|
|
|
|
|
Y.S.Jung,
M.Cai,
and
G.M.Clore
(2010).
Solution structure of the IIAChitobiose-IIBChitobiose complex of the N,N'-diacetylchitobiose branch of the Escherichia coli phosphotransferase system.
|
| |
J Biol Chem,
285,
4173-4184.
|
|
|
PDB codes:
|
|
|
|
|
|
|
|
G.M.Clore,
and
J.Iwahara
(2009).
Theory, practice, and applications of paramagnetic relaxation enhancement for the characterization of transient low-population states of biological macromolecules and their complexes.
|
| |
Chem Rev,
109,
4108-4139.
|
|
|
|
|
|
|
H.Feng,
L.M.Jenkins,
S.R.Durell,
R.Hayashi,
S.J.Mazur,
S.Cherry,
J.E.Tropea,
M.Miller,
A.Wlodawer,
E.Appella,
and
Y.Bai
(2009).
Structural basis for p300 Taz2-p53 TAD1 binding and modulation by phosphorylation.
|
| |
Structure,
17,
202-210.
|
|
|
PDB code:
|
|
|
|
|
|
|
|
D.J.Roy,
I.Casabon,
K.Vaillancourt,
J.L.Huot,
and
C.Vadeboncoeur
(2008).
Streptococci and lactococci synthesize large amounts of HPr(Ser-P)(His~P).
|
| |
Can J Microbiol,
54,
941-949.
|
|
|
|
|
|
|
J.Hu,
K.Hu,
D.C.Williams,
M.E.Komlosh,
M.Cai,
and
G.M.Clore
(2008).
Solution NMR structures of productive and non-productive complexes between the A and B domains of the cytoplasmic subunit of the mannose transporter of the Escherichia coli phosphotransferase system.
|
| |
J Biol Chem,
283,
11024-11037.
|
|
|
PDB codes:
|
|
|
|
|
|
|
|
J.Y.Suh,
M.Cai,
and
G.M.Clore
(2008).
Impact of phosphorylation on structure and thermodynamics of the interaction between the N-terminal domain of enzyme I and the histidine phosphocarrier protein of the bacterial phosphotransferase system.
|
| |
J Biol Chem,
283,
18980-18989.
|
|
|
|
|
|
|
Y.C.Kim,
C.Tang,
G.M.Clore,
and
G.Hummer
(2008).
Replica exchange simulations of transient encounter complexes in protein-protein association.
|
| |
Proc Natl Acad Sci U S A,
105,
12855-12860.
|
|
|
|
|
|
|
B.Reichenbach,
D.A.Breustedt,
J.Stülke,
B.Rak,
and
B.Görke
(2007).
Genetic dissection of specificity determinants in the interaction of HPr with enzymes II of the bacterial phosphoenolpyruvate:sugar phosphotransferase system in Escherichia coli.
|
| |
J Bacteriol,
189,
4603-4613.
|
|
|
|
|
|
|
J.Y.Suh,
J.Iwahara,
and
G.M.Clore
(2007).
Intramolecular domain-domain association/dissociation and phosphoryl transfer in the mannitol transporter of Escherichia coli are not coupled.
|
| |
Proc Natl Acad Sci U S A,
104,
3153-3158.
|
|
|
|
|
|
|
W.Müller,
and
H.Sticht
(2007).
A protein-specifically adapted scoring function for the reranking of docking solutions.
|
| |
Proteins,
67,
98.
|
|
|
|
|
|
|
C.Tang,
and
G.M.Clore
(2006).
A simple and reliable approach to docking protein-protein complexes from very sparse NOE-derived intermolecular distance restraints.
|
| |
J Biomol NMR,
36,
37-44.
|
|
|
|
|
|
|
C.Tang,
J.Iwahara,
and
G.M.Clore
(2006).
Visualization of transient encounter complexes in protein-protein association.
|
| |
Nature,
444,
383-386.
|
|
|
|
|
|
|
J.Deutscher,
C.Francke,
and
P.W.Postma
(2006).
How phosphotransferase system-related protein phosphorylation regulates carbohydrate metabolism in bacteria.
|
| |
Microbiol Mol Biol Rev,
70,
939.
|
|
|
|
|
|
|
J.Vaynberg,
and
J.Qin
(2006).
Weak protein-protein interactions as probed by NMR spectroscopy.
|
| |
Trends Biotechnol,
24,
22-27.
|
|
|
|
|
|
|
L.Volpon,
C.R.Young,
A.Matte,
and
K.Gehring
(2006).
NMR structure of the enzyme GatB of the galactitol-specific phosphoenolpyruvate-dependent phosphotransferase system and its interaction with GatA.
|
| |
Protein Sci,
15,
2435-2441.
|
|
|
PDB code:
|
|
|
|
|
|
|
|
D.C.Williams,
M.Cai,
J.Y.Suh,
A.Peterkofsky,
and
G.M.Clore
(2005).
Solution NMR structure of the 48-kDa IIAMannose-HPr complex of the Escherichia coli mannose phosphotransferase system.
|
| |
J Biol Chem,
280,
20775-20784.
|
|
|
PDB code:
|
|
|
|
|
|
|
|
J.Ren,
S.Sainsbury,
N.S.Berrow,
D.Alderton,
J.E.Nettleship,
D.K.Stammers,
N.J.Saunders,
and
R.J.Owens
(2005).
Crystal structure of nitrogen regulatory protein IIANtr from Neisseria meningitidis.
|
| |
BMC Struct Biol,
5,
13.
|
|
|
PDB code:
|
|
|
|
|
|
|
|
A.Mazé,
G.Boël,
S.Poncet,
I.Mijakovic,
Y.Le Breton,
A.Benachour,
V.Monedero,
J.Deutscher,
and
A.Hartke
(2004).
The Lactobacillus casei ptsHI47T mutation causes overexpression of a LevR-regulated but RpoN-independent operon encoding a mannose class phosphotransferase system.
|
| |
J Bacteriol,
186,
4543-4555.
|
|
|
|
|
|
|
Y.Qu,
J.T.Guo,
V.Olman,
and
Y.Xu
(2004).
Protein structure prediction using sparse dipolar coupling data.
|
| |
Nucleic Acids Res,
32,
551-561.
|
|
|
|
|
|
|
G.Wang,
P.A.Keifer,
and
A.Peterkofsky
(2003).
Solution structure of the N-terminal amphitropic domain of Escherichia coli glucose-specific enzyme IIA in membrane-mimetic micelles.
|
| |
Protein Sci,
12,
1087-1096.
|
|
|
PDB codes:
|
|
|
|
|
|
|
The most recent references are shown first.
Citation data come partly from CiteXplore and partly
from an automated harvesting procedure. Note that this is likely to be
only a partial list as not all journals are covered by
either method. However, we are continually building up the citation data
so more and more references will be included with time.
Where a reference describes a PDB structure, the PDB
codes are
shown on the right.
|
 |


 Links
Links















