 |
PDBsum entry 2k8f

|
|
|
|
|
|

|

|

|

|
|

|
|

|
|

|
|

|
|

|
|
|
 |

|

|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Transferase/transcription
|
PDB id
|
|
|
|
2k8f
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|

|

|

|
|
|
|
|
|
|
|
 |
Contents |
 |
|
|
|
|
|
|
|
|
* Residue conservation analysis
|
|
|
|
|
|
|
|
|
|
Enzyme class 1:
|
|
Chain A:
E.C.2.3.1.-
- ?????
|
|
|
|
|
|
|
Enzyme class 2:
|
|
Chain A:
E.C.2.3.1.48
- histone acetyltransferase.
|
|
|
|
|
|
|
Reaction:
|
|
L-lysyl-[protein] + acetyl-CoA = N6-acetyl-L-lysyl-[protein] + CoA + H+
|
|
|
|
|
|
L-lysyl-[protein]
|
+
|
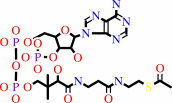 acetyl-CoA
acetyl-CoA
|
=
|
N(6)-acetyl-L-lysyl-[protein]
|
+
|
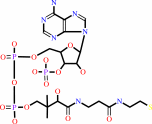 CoA
CoA
|
+
|
H(+)
|
|
|
|
|
|
|
|
|
|
Enzyme class 3:
|
|
Chain B:
E.C.?
|
|
|
|
|
|
|
|
|
|
Note, where more than one E.C. class is given (as above), each may
correspond to a different protein domain or, in the case of polyprotein
precursors, to a different mature protein.
|
|
|
|
Molecule diagrams generated from .mol files obtained from the
KEGG ftp site
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
| |
|
|
| |
|
DOI no:
|
Structure
17:202-210
(2009)
|
|
PubMed id:
|
|
|
|
|
|
| |
|
Structural basis for p300 Taz2-p53 TAD1 binding and modulation by phosphorylation.
|
|
H.Feng,
L.M.Jenkins,
S.R.Durell,
R.Hayashi,
S.J.Mazur,
S.Cherry,
J.E.Tropea,
M.Miller,
A.Wlodawer,
E.Appella,
Y.Bai.
|
|
|
|
|
| |
ABSTRACT
|
|
|
|
| |
|
|
Coactivators CREB-binding protein and p300 play important roles in mediating the
transcriptional activity of p53. Until now, however, no detailed structural
information has been available on how any of the domains of p300 interact with
p53. Here, we report the NMR structure of the complex of the Taz2 (C/H3) domain
of p300 and the N-terminal transactivation domain of p53. In the complex, p53
forms a short alpha helix and interacts with the Taz2 domain through an extended
surface. Mutational analyses demonstrate the importance of hydrophobic residues
for complex stabilization. Additionally, they suggest that the increased
affinity of Taz2 for p53(1-39) phosphorylated at Thr(18) is due in part to
electrostatic interactions of the phosphate with neighboring arginine residues
in Taz2. Thermodynamic experiments revealed the importance of hydrophobic
interactions in the complex of Taz2 with p53 phosphorylated at Ser(15) and
Thr(18).
|
|
|
|
|
|
| |
Selected figure(s)
|
|
|
|
| |
|
|

|
|

|
|
Figure 1.
Figure 1. Structure of the Taz2-p53[2–39] Complex
(A)
Stereo image of the overlay of ten lowest-energy NMR structures
of the complex between p53[15–27] (magenta) and the Taz2
domain of p300 (gray). The structures are superimposed on the
Cα traces.
(B) Plot of backbone amide ^15N-{^1H}
heteronuclear NOEs of p53[2–39].
(C) Cylinder model of
the average conformation of the complex. p53 is shown in magenta
and the helices of Taz2 are shown in blue (α1), lilac (α2),
orange (α3), and red (α4). Zinc ions in Taz2, modeled as green
spheres, were added according to zinc-coordination distances
into the known binding cage.
(D) Secondary chemical shift
difference of p53[2-39] in the complex (measured Cα chemical
shift − random coil value).
|
|
Figure 2.
Figure 2. Stabilizing Interactions between p53[2–39] and
Taz2
(A) Model of the Taz2-p53[2–39] complex, showing
residues that make hydrophobic contacts. Contacting residues are
labeled in white for Taz2 (gray surface representation) and red
for p53[2–39] (blue ribbon).
(B) Model of the
Taz2-p53[2–39] complex, colored by electrostatic potential
(red represents negative, blue indicates positive). Taz2 is
shown in a solid representation, and p53[2–39] as a mesh.
(C) Model of the Taz2-p53[2–39] complex, showing residues that
make electrostatic contacts. Contacting residues are labeled in
white for Taz2 (gray surface representation) and red for
p53[2–39] (blue ribbon).
|
|
|
|
|
|
| |
The above figures are
reprinted
by permission from Cell Press:
Structure
(2009,
17,
202-210)
copyright 2009.
|
|
| |
Figures were
selected
by an automated process.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
 |
 |
|
Literature references that cite this PDB file's key reference
|
|
|
| |
PubMed id
|
|
Reference
|
|
|
|
|
|
A.C.Joerger,
and
A.R.Fersht
(2010).
The tumor suppressor p53: from structures to drug discovery.
|
| |
Cold Spring Harb Perspect Biol,
2,
a000919.
|
|
|
|
|
|
|
E.Herbig,
L.Warfield,
L.Fish,
J.Fishburn,
B.A.Knutson,
B.Moorefield,
D.Pacheco,
and
S.Hahn
(2010).
Mechanism of Mediator recruitment by tandem Gcn4 activation domains and three Gal11 activator-binding domains.
|
| |
Mol Cell Biol,
30,
2376-2390.
|
|
|
|
|
|
|
M.L.Nelson,
H.S.Kang,
G.M.Lee,
A.G.Blaszczak,
D.K.Lau,
L.P.McIntosh,
and
B.J.Graves
(2010).
Ras signaling requires dynamic properties of Ets1 for phosphorylation-enhanced binding to coactivator CBP.
|
| |
Proc Natl Acad Sci U S A,
107,
10026-10031.
|
|
|
PDB code:
|
|
|
|
|
|
|
|
M.V.Poyurovsky,
C.Katz,
O.Laptenko,
R.Beckerman,
M.Lokshin,
J.Ahn,
I.J.Byeon,
R.Gabizon,
M.Mattia,
A.Zupnick,
L.M.Brown,
A.Friedler,
and
C.Prives
(2010).
The C terminus of p53 binds the N-terminal domain of MDM2.
|
| |
Nat Struct Mol Biol,
17,
982-989.
|
|
|
|
|
|
|
D.W.Meek,
and
C.W.Anderson
(2009).
Posttranslational modification of p53: cooperative integrators of function.
|
| |
Cold Spring Harb Perspect Biol,
1,
a000950.
|
|
|
|
|
|
|
D.W.Meek
(2009).
Tumour suppression by p53: a role for the DNA damage response?
|
| |
Nat Rev Cancer,
9,
714-723.
|
|
|
|
|
|
|
M.Miller,
Z.Dauter,
S.Cherry,
J.E.Tropea,
and
A.Wlodawer
(2009).
Structure of the Taz2 domain of p300: insights into ligand binding.
|
| |
Acta Crystallogr D Biol Crystallogr,
65,
1301-1308.
|
|
|
PDB code:
|
|
|
|
|
|
|
The most recent references are shown first.
Citation data come partly from CiteXplore and partly
from an automated harvesting procedure. Note that this is likely to be
only a partial list as not all journals are covered by
either method. However, we are continually building up the citation data
so more and more references will be included with time.
Where a reference describes a PDB structure, the PDB
code is
shown on the right.
|
|
|


 Links
Links























