 |
PDBsum entry 2qy2

|
|
|
|
|
|

|

|

|

|
|

|
|

|
|

|
|

|
|

|
|
|
 |

|

|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Hydrolase, viral protein
|
PDB id
|
|
|
|
2qy2
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|

|

|

|
|
|
|
|
|
|
|
|
Enzyme class 2:
|
|
E.C.2.1.1.56
- mRNA (guanine-N(7))-methyltransferase.
|
|
|
|
|
|
|
Reaction:
|
|
a 5'-end (5'-triphosphoguanosine)-ribonucleoside in mRNA + S-adenosyl-L- methionine = a 5'-end (N(7)-methyl 5'-triphosphoguanosine)-ribonucleoside in mRNA + S-adenosyl-L-homocysteine
|
|
|
|
|
|
5'-end (5'-triphosphoguanosine)-ribonucleoside in mRNA
|
+
|
 S-adenosyl-L- methionine
S-adenosyl-L- methionine
|
=
|
5'-end (N(7)-methyl 5'-triphosphoguanosine)-ribonucleoside in mRNA
|
+
|
 S-adenosyl-L-homocysteine
S-adenosyl-L-homocysteine
|
|
|
|
|
|
|
|
|
|
Enzyme class 3:
|
|
E.C.2.7.7.50
- mRNA guanylyltransferase.
|
|
|
|
|
|
|
Reaction:
|
|
a 5'-end diphospho-ribonucleoside in mRNA + GTP + H+ = a 5'-end (5'-triphosphoguanosine)-ribonucleoside in mRNA + diphosphate
|
|
|
|
|
|
5'-end diphospho-ribonucleoside in mRNA
|
+
|
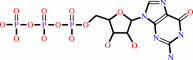 GTP
GTP
|
+
|
H(+)
|
=
|
5'-end (5'-triphosphoguanosine)-ribonucleoside in mRNA
|
+
|
 diphosphate
diphosphate
|
|
|
|
|
|
|
|
|
|
Enzyme class 4:
|
|
E.C.3.6.1.74
- mRNA 5'-phosphatase.
|
|
|
|
|
|
|
Reaction:
|
|
a 5'-end triphospho-ribonucleoside in mRNA + H2O = a 5'-end diphospho- ribonucleoside in mRNA + phosphate + H+
|
|
|
|
|
|
5'-end triphospho-ribonucleoside in mRNA
|
+
|
H2O
|
=
|
5'-end diphospho- ribonucleoside in mRNA
|
+
|
 phosphate
phosphate
|
+
|
H(+)
|
|
|
|
|
|
|
|
|
|
|
|
|
Note, where more than one E.C. class is given (as above), each may
correspond to a different protein domain or, in the case of polyprotein
precursors, to a different mature protein.
|
|
|
|
Molecule diagrams generated from .mol files obtained from the
KEGG ftp site
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
| |
|
|
| |
|
DOI no:
|
Structure
16:501-512
(2008)
|
|
PubMed id:
|
|
|
|
|
|
| |
|
Characterization of a trifunctional mimivirus mRNA capping enzyme and crystal structure of the RNA triphosphatase domain.
|
|
D.Benarroch,
P.Smith,
S.Shuman.
|
|
|
|
|
| |
ABSTRACT
|
|
|
|
| |
|
|
The RNA triphosphatase (RTPase) components of the mRNA capping apparatus are a
bellwether of eukaryal taxonomy. Fungal and protozoal RTPases belong to the
triphosphate tunnel metalloenzyme (TTM) family, exemplified by yeast Cet1.
Several large DNA viruses encode metal-dependent RTPases unrelated to the
cysteinyl-phosphatase RTPases of their metazoan host organisms. The origins of
DNA virus RTPases are unclear because they are structurally uncharacterized.
Mimivirus, a giant virus of amoeba, resembles poxviruses in having a
trifunctional capping enzyme composed of a metal-dependent RTPase module fused
to guanylyltransferase (GTase) and guanine-N7 methyltransferase domains. The
crystal structure of mimivirus RTPase reveals a minimized tunnel fold and an
active site strikingly similar to that of Cet1. Unlike homodimeric fungal
RTPases, mimivirus RTPase is a monomer. The mimivirus TTM-type RTPase-GTase
fusion resembles the capping enzymes of amoebae, providing evidence that the
ancestral large DNA virus acquired its capping enzyme from a unicellular host.
|
|
|
|
|
|
| |
Selected figure(s)
|
|
|
|
| |
|
|

|
|

|
|
Figure 5.
Figure 5. Tertiary Structure of Mimivirus RTPase
(A–F)
The fold of mimivirus RTPase (amino acids 11–237) is depicted
as a ribbon diagram in (A), (C), and (D); α helices are colored
cyan, and β strands are colored magenta. A view into the
triphosphate tunnel is highlighted in (A). The N and C termini
are indicated. The images in (C) and (D) are rotated clockwise
and counterclockwise, respectively, with respect to (A) in order
to highlight side views of the staves of the β barrel. (B)
shows a space-filling surface model in the same orientation as
(A) that highlights the tunnel aperture and an acetate molecule
(depicted as a stick model) in the center of the tunnel. The
primary structure is displayed in (E); secondary structure
elements are highlighted in cyan for α helices and magenta for
β strands. The putative metal-binding motifs are located in
strands β1 and β8; the essential glutamates are denoted by
dots (•). The ^121DIEIVYKN^128 and ^133KLIGI^137 β segments
that are interrupted by a short non-β ^129RGSG^132 peptide
(indicated by an asterisk in [A]) together comprise one of the
barrel staves, which will be considered as a single β element
(indicated by brackets in [E]) that corresponds to the fourth β
strand of the triphosphate tunnel of yeast Cet1. (F) shows a
comparison of the topologies of mimivirus RTPase and yeast Cet1.
Tunnel β strands are shown as magenta pentagons oriented in the
flat plane according to the view in (A), such that pentagons
with the apices pointing into the tunnel in (F) correspond to
strands that project out from the page toward the viewer in (A),
while pentagons with apices pointing away from the tunnel in (F)
are ones that project into the plane of the page in (A). The
mimivirus RTPase α helices are shown as cyan circles, as are
the corresponding α helices in Cet1. Additional secondary
structure elements unique to Cet1 are colored gray. A disordered
chain break on the tunnel roof of mimivirus RTPase (from amino
acid 155 to amino acid 157 in the loop connecting strands 5 and
6) is indicated by a dashed line. This segment is ordered in the
A protomer of the monoclinic crystal, as a result of crystal
packing contacts unique to the monoclinic lattice. Chains breaks
occurring at different sites in Cet1 are denoted by dashed lines.
|
|
Figure 6.
Figure 6. MimiCE RTPase Active Site and Comparison to Cet1
(A and B) Stereo views of the tunnel interiors of (A)
MimiCE-(1–237) and (B) yeast Cet1. Waters are depicted as red
spheres. The Cet1-bound manganese ion is a cyan sphere. Acetate
and sulfate ions in the tunnels are rendered as stick models, as
are side chains emanating from the β strands that comprise the
tunnel walls.
|
|
|
|
|
|
| |
The above figures are
reprinted
by permission from Cell Press:
Structure
(2008,
16,
501-512)
copyright 2008.
|
|
| |
Figures were
selected
by an automated process.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
 |
 |
|
Literature references that cite this PDB file's key reference
|
|
|
| |
PubMed id
|
|
Reference
|
|
|
|
|
|
A.Simoes-Barbosa,
R.P.Hirt,
and
P.J.Johnson
(2010).
A metazoan/plant-like capping enzyme and cap modified nucleotides in the unicellular eukaryote Trichomonas vaginalis.
|
| |
PLoS Pathog,
6,
e1000999.
|
|
|
|
|
|
|
M.Gu,
K.R.Rajashankar,
and
C.D.Lima
(2010).
Structure of the Saccharomyces cerevisiae Cet1-Ceg1 mRNA capping apparatus.
|
| |
Structure,
18,
216-227.
|
|
|
PDB code:
|
|
|
|
|
|
|
|
P.A.Nair,
P.Smith,
and
S.Shuman
(2010).
Structure of bacterial LigD 3'-phosphoesterase unveils a DNA repair superfamily.
|
| |
Proc Natl Acad Sci U S A,
107,
12822-12827.
|
|
|
PDB codes:
|
|
|
|
|
|
|
|
D.Benarroch,
Z.R.Qiu,
B.Schwer,
and
S.Shuman
(2009).
Characterization of a mimivirus RNA cap guanine-N2 methyltransferase.
|
| |
RNA,
15,
666-674.
|
|
|
|
|
|
|
D.Byrne,
R.Grzela,
A.Lartigue,
S.Audic,
S.Chenivesse,
S.Encinas,
J.M.Claverie,
and
C.Abergel
(2009).
The polyadenylation site of Mimivirus transcripts obeys a stringent 'hairpin rule'.
|
| |
Genome Res,
19,
1233-1242.
|
|
|
|
|
|
|
J.M.Claverie,
and
C.Abergel
(2009).
Mimivirus and its virophage.
|
| |
Annu Rev Genet,
43,
49-66.
|
|
|
|
|
|
|
M.Issur,
S.Despins,
I.Bougie,
and
M.Bisaillon
(2009).
Nucleotide analogs and molecular modeling studies reveal key interactions involved in substrate recognition by the yeast RNA triphosphatase.
|
| |
Nucleic Acids Res,
37,
3714-3722.
|
|
|
|
|
|
|
R.Jain,
and
S.Shuman
(2008).
Polyphosphatase Activity of CthTTM, a Bacterial Triphosphate Tunnel Metalloenzyme.
|
| |
J Biol Chem,
283,
31047-31057.
|
|
|
|
|
|
The most recent references are shown first.
Citation data come partly from CiteXplore and partly
from an automated harvesting procedure. Note that this is likely to be
only a partial list as not all journals are covered by
either method. However, we are continually building up the citation data
so more and more references will be included with time.
Where a reference describes a PDB structure, the PDB
code is
shown on the right.
|
|


 Links
Links















