 |
PDBsum entry 1foh

|
|
|
|
|
 |
Contents |
 |
|
|
|
|
|
|
|
|
|
|
|
|
* Residue conservation analysis
|
|
|
|
|
|
|
|
|
|
|
Enzyme class:
|
|
E.C.1.14.13.7
- phenol 2-monooxygenase (NADPH).
|
|
|
|
|
|
|
Reaction:
|
|
phenol + NADPH + O2 + H+ = catechol + NADP+ + H2O
|
|
|
|
|
|
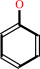 phenol
phenol
|
+
|
 NADPH
NADPH
|
+
|
 O2
O2
|
+
|
H(+)
Bound ligand (Het Group name = )
corresponds exactly
|
=
|
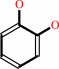 catechol
catechol
|
+
|
 NADP(+)
NADP(+)
|
+
|
H2O
|
|
|
|
|
|
|
|
|
|
Cofactor:
|
|
FAD
|
|
|
|
|
|
FAD
Bound ligand (Het Group name =
FAD)
corresponds exactly
|
|
|
|
|
|
|
Molecule diagrams generated from .mol files obtained from the
KEGG ftp site
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
| |
|
|
| |
|
DOI no:
|
Structure
6:605-617
(1998)
|
|
PubMed id:
|
|
|
|
|
|
| |
|
The crystal structure of phenol hydroxylase in complex with FAD and phenol provides evidence for a concerted conformational change in the enzyme and its cofactor during catalysis.
|
|
C.Enroth,
H.Neujahr,
G.Schneider,
Y.Lindqvist.
|
|
|
|
|
| |
ABSTRACT
|
|
|
|
| |
|
|
BACKGROUND: The synthesis of phenolic compounds as by-products of industrial
reactions poses a serious threat to the environment. Understanding the enzymatic
reactions involved in the degradation and detoxification of these compounds is
therefore of much interest. Soil-living yeasts use flavin adenine dinucleotide
(FAD)-containing enzymes to hydroxylate phenols. This reaction initiates a
metabolic sequence permitting utilisation of the aromatic compound as a source
of carbon and energy. The phenol hydroxylase from Trichosporon cutaneum
hydroxylates phenol to catechol. Phenol is the best substrate, but the enzyme
also accepts simple hydroxyl-, amino-, halogen- or methyl-substituted phenols.
RESULTS: The crystal structure of phenol hydroxylase in complex with FAD and
phenol has been determined at 2.4 A resolution. The structure was solved by the
MIRAS method. The protein model consists of two homodimers. The subunit consists
of three domains, the first of which contains a beta sheet that binds FAD with a
typical beta alpha beta nucleotide-binding motif and also a fingerprint motif
for NADPH binding. The active site is located at the interface between the first
and second domains; the second domain also binds the phenolic substrate. The
third domain contains a thioredoxin-like fold and is involved in dimer contacts.
The subunits within the dimer show substantial differences in structure and in
FAD conformation. This conformational flexibility allows the substrate to gain
access to the active site and excludes solvent during the hydroxylation
reaction. CONCLUSIONS: Two of the domains of phenol hydroxylase are similar in
structure to p-hydroxybenzoate hydroxylase. Thus, phenol hydroxylase is a member
of a family of flavin-containing aromatic hydroxylases that share the same
overall fold, in spite of large differences in amino acid sequences and chain
length. The structure of phenol hydroxylase is consistent with a hydroxyl
transfer mechanism via a peroxo-FAD intermediate. We propose that a movement of
FAD takes place in concert with a large conformational change of residues
170-210 during catalysis.
|
|
|
|
|
|
| |
Selected figure(s)
|
|
|
|
| |
|
|

|
|
Figure 6.
Figure 6. Schematic drawing of the FAD-binding site of
phenol hydroxylase in (a) the `in' conformation and (b) the
`out' conformation. The two conformations are stabilised by
hydrogen bonds, but the bonding pattern is significantly
different in the two states. Most notable are the hydrogen bonds
from the isoalloxazine ring to the mainchain nitrogen atoms of
residues Gly55, Gly369, Met370 and Asn371, which are all broken
and in some cases exchanged to hydrogen bonds to water molecules
in the out conformation. Residue His 189 (in (a)), depicted with
an asterisk, is located in the flexible segment of the
polypeptide chain and consequently does not make contacts to FAD
in the out conformation, where the loop is in the open
conformation. In addition, residues 43-52 move slightly between
the two conformations, in concert with the movement of the
isoalloxazine ring. Most notable here is Gln52, which works as a
hinge for this stretch of residues. In the in case Gln52 is
hydrogen bonded to an oxygen atom of the ribityl chain; in the
out case Gln52 has moved its sidechain to lie parallel to the
isoalloxazine ring. A new hydrogen bond is formed from the same
oxygen, but this time to Gln117. Most of the hydrogen bonds to
FAD are conserved for this class of aromatic hydroxylases.
|
|
|
|
|
|
| |
The above figure is
reprinted
by permission from Cell Press:
Structure
(1998,
6,
605-617)
copyright 1998.
|
|
| |
Figure was
selected
by an automated process.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
 |
 |
|
Literature references that cite this PDB file's key reference
|
|
|
| |
PubMed id
|
|
Reference
|
|
|
|
|
|
K.Groom,
A.Bhattacharya,
and
D.L.Zechel
(2011).
Rebeccamycin and Staurosporine Biosynthesis: Insight into the Mechanisms of the Flavin-Dependent Monooxygenases RebC and StaC.
|
| |
Chembiochem,
12,
396-400.
|
|
|
|
|
|
|
S.Sah,
and
P.S.Phale
(2011).
1-Naphthol 2-hydroxylase from Pseudomonas sp. strain C6: purification, characterization and chemical modification studies.
|
| |
Biodegradation,
22,
517-526.
|
|
|
|
|
|
|
T.Awakawa,
N.Fujita,
M.Hayakawa,
Y.Ohnishi,
and
S.Horinouchi
(2011).
Characterization of the Biosynthesis Gene Cluster for Alkyl-O-Dihydrogeranyl-Methoxyhydroquinones in Actinoplanes missouriensis.
|
| |
Chembiochem,
12,
439-448.
|
|
|
|
|
|
|
B.Tian,
Y.Tu,
A.Strid,
and
L.A.Eriksson
(2010).
Hydroxylation and ring-opening mechanism of an unusual flavoprotein monooxygenase, 2-methyl-3-hydroxypyridine-5-carboxylic acid oxygenase: a theoretical study.
|
| |
Chemistry,
16,
2557-2566.
|
|
|
|
|
|
|
U.E.Ukaegbu,
A.Kantz,
M.Beaton,
G.T.Gassner,
and
A.C.Rosenzweig
(2010).
Structure and ligand binding properties of the epoxidase component of styrene monooxygenase .
|
| |
Biochemistry,
49,
1678-1688.
|
|
|
PDB code:
|
|
|
|
|
|
|
|
J.J.Zhang,
H.Liu,
Y.Xiao,
X.E.Zhang,
and
N.Y.Zhou
(2009).
Identification and characterization of catabolic para-nitrophenol 4-monooxygenase and para-benzoquinone reductase from Pseudomonas sp. strain WBC-3.
|
| |
J Bacteriol,
191,
2703-2710.
|
|
|
|
|
|
|
K.M.Meneely,
E.W.Barr,
J.M.Bollinger,
and
A.L.Lamb
(2009).
Kinetic mechanism of ornithine hydroxylase (PvdA) from Pseudomonas aeruginosa: substrate triggering of O2 addition but not flavin reduction.
|
| |
Biochemistry,
48,
4371-4376.
|
|
|
|
|
|
|
W.Tong,
Y.Wei,
L.F.Murga,
M.J.Ondrechen,
and
R.J.Williams
(2009).
Partial order optimum likelihood (POOL): maximum likelihood prediction of protein active site residues using 3D Structure and sequence properties.
|
| |
PLoS Comput Biol,
5,
e1000266.
|
|
|
|
|
|
|
J.Xiao,
and
J.M.Vanbriesen
(2008).
Expanded thermodynamic true yield prediction model: adjustments and limitations.
|
| |
Biodegradation,
19,
99.
|
|
|
|
|
|
|
M.Funabashi,
N.Funa,
and
S.Horinouchi
(2008).
Phenolic lipids synthesized by type III polyketide synthase confer penicillin resistance on Streptomyces griseus.
|
| |
J Biol Chem,
283,
13983-13991.
|
|
|
|
|
|
|
K.Palmu,
K.Ishida,
P.Mäntsälä,
C.Hertweck,
and
M.Metsä-Ketelä
(2007).
Artificial reconstruction of two cryptic angucycline antibiotic biosynthetic pathways.
|
| |
Chembiochem,
8,
1577-1584.
|
|
|
|
|
|
|
M.Zhao,
X.Jia,
C.Wang,
Q.Li,
K.Zhou,
L.Wang,
H.Liu,
and
S.Peng
(2007).
PAK: an essential motif for forming beta-turn structures and exhibiting the thrombolytic effect of P6A and its analogs.
|
| |
Biochem Cell Biol,
85,
730-740.
|
|
|
|
|
|
|
S.H.Kim,
T.Hisano,
K.Takeda,
W.Iwasaki,
A.Ebihara,
and
K.Miki
(2007).
Crystal structure of the oxygenase component (HpaB) of the 4-hydroxyphenylacetate 3-monooxygenase from Thermus thermophilus HB8.
|
| |
J Biol Chem,
282,
33107-33117.
|
|
|
PDB codes:
|
|
|
|
|
|
|
|
T.N.Gustafsson,
T.Sandalova,
J.Lu,
A.Holmgren,
and
G.Schneider
(2007).
High-resolution structures of oxidized and reduced thioredoxin reductase from Helicobacter pylori.
|
| |
Acta Crystallogr D Biol Crystallogr,
63,
833-843.
|
|
|
PDB codes:
|
|
|
|
|
|
|
|
K.A.Feenstra,
K.Hofstetter,
R.Bosch,
A.Schmid,
J.N.Commandeur,
and
N.P.Vermeulen
(2006).
Enantioselective substrate binding in a monooxygenase protein model by molecular dynamics and docking.
|
| |
Biophys J,
91,
3206-3216.
|
|
|
PDB code:
|
|
|
|
|
|
|
|
M.Nadella,
M.A.Bianchet,
S.B.Gabelli,
J.Barrila,
and
L.M.Amzel
(2005).
Structure and activity of the axon guidance protein MICAL.
|
| |
Proc Natl Acad Sci U S A,
102,
16830-16835.
|
|
|
PDB code:
|
|
|
|
|
|
|
|
W.Oonanant,
J.Sucharitakul,
J.Yuvaniyama,
and
P.Chaiyen
(2005).
Crystallization and preliminary X-ray crystallographic analysis of 2-methyl-3-hydroxypyridine-5-carboxylic acid (MHPC) oxygenase from Pseudomonas sp. MA-1.
|
| |
Acta Crystallogr Sect F Struct Biol Cryst Commun,
61,
312-314.
|
|
|
|
|
|
|
M.Dai,
and
S.D.Copley
(2004).
Genome shuffling improves degradation of the anthropogenic pesticide pentachlorophenol by Sphingobium chlorophenolicum ATCC 39723.
|
| |
Appl Environ Microbiol,
70,
2391-2397.
|
|
|
|
|
|
|
A.Meyer,
D.Tanner,
A.Schmid,
D.F.Sargent,
H.P.Kohler,
and
B.Witholt
(2003).
Crystallization and preliminary X-ray analysis of native and selenomethionine 2-hydroxybiphenyl 3-monooxygenase.
|
| |
Acta Crystallogr D Biol Crystallogr,
59,
741-743.
|
|
|
|
|
|
|
C.Enroth
(2003).
High-resolution structure of phenol hydroxylase and correction of sequence errors.
|
| |
Acta Crystallogr D Biol Crystallogr,
59,
1597-1602.
|
|
|
PDB code:
|
|
|
|
|
|
|
|
T.Nakamura,
T.Motoyama,
T.Hirokawa,
S.Hirono,
and
I.Yamaguchi
(2003).
Computer-aided modeling of pentachlorophenol 4-monooxygenase and site-directed mutagenesis of its active site.
|
| |
Chem Pharm Bull (Tokyo),
51,
1293-1298.
|
|
|
|
|
|
|
U.Kirchner,
A.H.Westphal,
R.Müller,
and
W.J.van Berkel
(2003).
Phenol hydroxylase from Bacillus thermoglucosidasius A7, a two-protein component monooxygenase with a dual role for FAD.
|
| |
J Biol Chem,
278,
47545-47553.
|
|
|
|
|
|
|
A.Meyer,
A.Schmid,
M.Held,
A.H.Westphal,
M.Rothlisberger,
H.P.Kohler,
W.J.van Berkel,
and
B.Witholt
(2002).
Changing the substrate reactivity of 2-hydroxybiphenyl 3-monooxygenase from Pseudomonas azelaica HBP1 by directed evolution.
|
| |
J Biol Chem,
277,
5575-5582.
|
|
|
|
|
|
|
A.Meyer,
M.Würsten,
A.Schmid,
H.P.Kohler,
and
B.Witholt
(2002).
Hydroxylation of indole by laboratory-evolved 2-hydroxybiphenyl 3-monooxygenase.
|
| |
J Biol Chem,
277,
34161-34167.
|
|
|
|
|
|
|
B.A.Palfey,
R.Basu,
K.K.Frederick,
B.Entsch,
and
D.P.Ballou
(2002).
Role of protein flexibility in the catalytic cycle of p-hydroxybenzoate hydroxylase elucidated by the Pro293Ser mutant.
|
| |
Biochemistry,
41,
8438-8446.
|
|
|
|
|
|
|
M.D.Altose,
Y.Zheng,
J.Dong,
B.A.Palfey,
and
P.R.Carey
(2001).
Comparing protein-ligand interactions in solution and single crystals by Raman spectroscopy.
|
| |
Proc Natl Acad Sci U S A,
98,
3006-3011.
|
|
|
|
|
|
|
M.D.Krasowski,
K.Nishikawa,
N.Nikolaeva,
A.Lin,
and
N.L.Harrison
(2001).
Methionine 286 in transmembrane domain 3 of the GABAA receptor beta subunit controls a binding cavity for propofol and other alkylphenol general anesthetics.
|
| |
Neuropharmacology,
41,
952-964.
|
|
|
|
|
|
|
M.Ortiz-Maldonado,
D.P.Ballou,
and
V.Massey
(2001).
A rate-limiting conformational change of the flavin in p-hydroxybenzoate hydroxylase is necessary for ligand exchange and catalysis: studies with 8-mercapto- and 8-hydroxy-flavins.
|
| |
Biochemistry,
40,
1091-1101.
|
|
|
|
|
|
|
O.Dym,
and
D.Eisenberg
(2001).
Sequence-structure analysis of FAD-containing proteins.
|
| |
Protein Sci,
10,
1712-1728.
|
|
|
|
|
|
|
J.Beynon,
E.R.Rafanan,
B.Shen,
and
A.J.Fisher
(2000).
Crystallization and preliminary X-ray analysis of tetracenomycin A2 oxygenase: a flavoprotein hydroxylase involved in polyketide biosynthesis.
|
| |
Acta Crystallogr D Biol Crystallogr,
56,
1647-1651.
|
|
|
|
|
|
|
M.H.Eppink,
E.Cammaart,
D.Van Wassenaar,
W.J.Middelhoven,
and
W.J.van Berkel
(2000).
Purification and properties of hydroquinone hydroxylase, a FAD-dependent monooxygenase involved in the catabolism of 4-hydroxybenzoate in Candida parapsilosis CBS604.
|
| |
Eur J Biochem,
267,
6832-6840.
|
|
|
|
|
|
|
O.Vallon
(2000).
New sequence motifs in flavoproteins: evidence for common ancestry and tools to predict structure.
|
| |
Proteins,
38,
95.
|
|
|
|
|
|
|
M.Ortiz-Maldonado,
D.Gatti,
D.P.Ballou,
and
V.Massey
(1999).
Structure-function correlations of the reaction of reduced nicotinamide analogues with p-hydroxybenzoate hydroxylase substituted with a series of 8-substituted flavins.
|
| |
Biochemistry,
38,
16636-16647.
|
|
|
PDB code:
|
|
|
|
|
|
|
|
M.Stehr,
L.Smau,
M.Singh,
O.Seth,
P.Macheroux,
S.Ghisla,
and
H.Diekmann
(1999).
Studies with lysine N6-hydroxylase. Effect of a mutation in the assumed FAD binding site on coenzyme affinities and on lysine hydroxylating activity.
|
| |
Biol Chem,
380,
47-54.
|
|
|
|
|
|
|
P.Trickey,
M.A.Wagner,
M.S.Jorns,
and
F.S.Mathews
(1999).
Monomeric sarcosine oxidase: structure of a covalently flavinylated amine oxidizing enzyme.
|
| |
Structure,
7,
331-345.
|
|
|
PDB codes:
|
|
|
|
|
|
|
|
W.A.Suske,
W.J.van Berkel,
and
H.P.Kohler
(1999).
Catalytic mechanism of 2-hydroxybiphenyl 3-monooxygenase, a flavoprotein from Pseudomonas azelaica HBP1.
|
| |
J Biol Chem,
274,
33355-33365.
|
|
|
|
|
|
|
Y.Zheng,
J.Dong,
B.A.Palfey,
and
P.R.Carey
(1999).
Using Raman spectroscopy to monitor the solvent-exposed and "buried" forms of flavin in p-hydroxybenzoate hydroxylase.
|
| |
Biochemistry,
38,
16727-16732.
|
|
|
|
|
|
The most recent references are shown first.
Citation data come partly from CiteXplore and partly
from an automated harvesting procedure. Note that this is likely to be
only a partial list as not all journals are covered by
either method. However, we are continually building up the citation data
so more and more references will be included with time.
Where a reference describes a PDB structure, the PDB
code is
shown on the right.
|
|


 Links
Links














