 |
PDBsum entry 5ezs

|
|
|
|
|
|

|

|

|

|
|

|
|

|
|

|
|
|
 |

|

|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Hydrolase/hydrolase inhibitor
|
PDB id
|
|
|
|
5ezs
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|

|

|

|
|
|
|
|
|
|
|
|
Enzyme class 1:
|
|
E.C.2.1.1.-
- ?????
|
|
|
|
|
|
|
Enzyme class 2:
|
|
E.C.2.7.7.-
- ?????
|
|
|
|
|
|
|
Enzyme class 3:
|
|
E.C.2.7.7.19
- polynucleotide adenylyltransferase.
|
|
|
|
|
|
|
Reaction:
|
|
RNA(n) + ATP = RNA(n)-3'-adenine ribonucleotide + diphosphate
|
|
|
|
|
|
 RNA(n)
RNA(n)
|
+
|
 ATP
ATP
|
=
|
RNA(n)-3'-adenine ribonucleotide
|
+
|
 diphosphate
diphosphate
|
|
|
|
|
|
|
|
|
|
Enzyme class 4:
|
|
E.C.2.7.7.48
- RNA-directed Rna polymerase.
|
|
|
|
|
|
|
Reaction:
|
|
RNA(n) + a ribonucleoside 5'-triphosphate = RNA(n+1) + diphosphate
|
|
|
|
|
|
RNA(n)
|
+
|
ribonucleoside 5'-triphosphate
|
=
|
RNA(n+1)
|
+
|
diphosphate
|
|
|
|
|
|
|
|
|
|
Enzyme class 5:
|
|
E.C.3.1.3.84
- ADP-ribose 1''-phosphate phosphatase.
|
|
|
|
|
|
|
Reaction:
|
|
ADP-alpha-D-ribose 1''-phosphate + H2O = ADP-D-ribose + phosphate
|
|
|
|
|
|
ADP-alpha-D-ribose 1''-phosphate
|
+
|
H2O
|
=
|
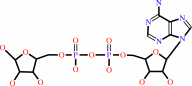 ADP-D-ribose
ADP-D-ribose
|
+
|
 phosphate
phosphate
|
|
|
|
|
|
|
|
|
|
Enzyme class 6:
|
|
E.C.3.4.22.-
- ?????
|
|
|
|
|
|
|
Enzyme class 7:
|
|
E.C.3.6.1.15
- nucleoside-triphosphate phosphatase.
|
|
|
|
|
|
|
Reaction:
|
|
a ribonucleoside 5'-triphosphate + H2O = a ribonucleoside 5'-diphosphate + phosphate + H+
|
|
|
|
|
|
ribonucleoside 5'-triphosphate
|
+
|
H2O
|
=
|
ribonucleoside 5'-diphosphate
|
+
|
phosphate
|
+
|
H(+)
|
|
|
|
|
|
|
|
|
|
Enzyme class 8:
|
|
E.C.3.6.1.74
- mRNA 5'-phosphatase.
|
|
|
|
|
|
|
Reaction:
|
|
a 5'-end triphospho-ribonucleoside in mRNA + H2O = a 5'-end diphospho- ribonucleoside in mRNA + phosphate + H+
|
|
|
|
|
|
5'-end triphospho-ribonucleoside in mRNA
|
+
|
H2O
|
=
|
5'-end diphospho- ribonucleoside in mRNA
|
+
|
phosphate
|
+
|
H(+)
|
|
|
|
|
|
|
|
|
|
Enzyme class 9:
|
|
E.C.3.6.4.13
- Rna helicase.
|
|
|
|
|
|
|
Reaction:
|
|
ATP + H2O = ADP + phosphate + H+
|
|
|
|
|
|
ATP
|
+
|
H2O
|
=
|
 ADP
ADP
|
+
|
phosphate
|
+
|
H(+)
|
|
|
|
|
|
|
|
|
|
|
|
|
Note, where more than one E.C. class is given (as above), each may
correspond to a different protein domain or, in the case of polyprotein
precursors, to a different mature protein.
|
|
|
|
Molecule diagrams generated from .mol files obtained from the
KEGG ftp site
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
| |
|
|
| |
|
DOI no:
|
Biochemistry
55:3007-3019
(2016)
|
|
PubMed id:
|
|
|
|
|
|
| |
|
Kinetic, Mutational, and Structural Studies of the Venezuelan Equine Encephalitis Virus Nonstructural Protein 2 Cysteine Protease.
|
|
X.Hu,
J.R.Compton,
D.H.Leary,
M.A.Olson,
M.S.Lee,
J.Cheung,
W.Ye,
M.Ferrer,
N.Southall,
A.Jadhav,
E.M.Morazzani,
P.J.Glass,
J.Marugan,
P.M.Legler.
|
|
|
|
|
| |
ABSTRACT
|
|
|
|
| |
|
|
The Venezuelan equine encephalitis virus (VEEV) nonstructural protein 2 (nsP2)
cysteine protease (EC 3.4.22.-) is essential for viral replication and is
involved in the cytopathic effects (CPE) of the virus. The VEEV nsP2 protease is
a member of MEROPS Clan CN and characteristically contains a papain-like
protease linked to an S-adenosyl-l-methionine-dependent RNA methyltransferase
(SAM MTase) domain. The protease contains an alternative active site motif,
(475)NVCWAK(480), which differs from papain's (CGS(25)CWAFS), and the enzyme
lacks a transition state-stabilizing residue homologous to Gln-19 in papain. To
understand the roles of conserved residues in catalysis, we determined the
structure of the free enzyme and the first structure of an inhibitor-bound
alphaviral protease. The peptide-like E64d inhibitor was found to bind beneath a
β-hairpin at the interface of the SAM MTase and protease domains. His-546
adopted a conformation that differed from that found in the free enzyme; one or
both of the conformers may assist in leaving group departure of either the amine
or Cys thiolate during the catalytic cycle. Interestingly, E64c (200 μM), the
carboxylic acid form of the E64d ester, did not inhibit the nsP2 protease. To
identify key residues involved in substrate binding, a number of mutants were
analyzed. Mutation of the motif residue, N475A, led to a 24-fold reduction in
kcat/Km, and the conformation of this residue did not change after inhibition.
N475 forms a hydrogen bond with R662 in the SAM MTase domain, and the R662A and
R662K mutations both led to 16-fold decreases in kcat/Km. N475 forms the base of
the P1 binding site and likely orients the substrate for nucleophilic attack or
plays a role in product release. An Asn homologous to N475 is similarly found in
coronaviral papain-like proteases (PLpro) of the Severe Acute Respiratory
Syndrome (SARS) virus and Middle East Respiratory Syndrome (MERS) virus.
Mutation of another motif residue, K480A, led to a 9-fold decrease in kcat and
kcat/Km. K480 likely enhances the nucleophilicity of the Cys. Consistent with
our substrate-bound models, the SAM MTase domain K706A mutation increased Km
4.5-fold to 500 μM. Within the β-hairpin, the N545A mutation slightly but not
significantly increased kcat and Km. The structures and identified active site
residues may facilitate the discovery of protease inhibitors with antiviral
activity.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
 |
 |
|


 Links
Links



















