
|
|
|
|
|
 |
Contents |
 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
* Residue conservation analysis
|
|
|
|
|
|
PDB id:
|
|
|
|
| Name: |
|
Oxidoreductase
|
|
|
Title:
|
|
Branched-chain 2-oxo acid dehydrogenase (e1) from thermus thermophilus hb8 with 4-methylpentanoate
|
|
Structure:
|
|
2-oxo acid dehydrogenase alpha subunit. Chain: a, c. Synonym: e1-alpha. Engineered: yes. 2-oxo acid dehydrogenase beta subunit. Chain: b, d. Synonym: e1-beta. Engineered: yes
|
|
Source:
|
|
Thermus thermophilus. Organism_taxid: 274. Expressed in: escherichia coli. Expression_system_taxid: 562.
|
|
Biol. unit:
|
|
Tetramer (from
)
|
|
Resolution:
|
|
|
2.40Å
|
R-factor:
|
0.172
|
R-free:
|
0.206
|
|
|
Authors:
|
|
T.Nakai,N.Nakagawa,N.Maoka,R.Masui,S.Kuramitsu,N.Kamiya,Riken Structural Genomics/proteomics Initiative (Rsgi)
|
Key ref:

|
|
T.Nakai
et al.
(2004).
Ligand-induced conformational changes and a reaction intermediate in branched-chain 2-oxo acid dehydrogenase (E1) from Thermus thermophilus HB8, as revealed by X-ray crystallography.
J Mol Biol,
337,
1011-1033.
PubMed id:
DOI:

|
|
|
Date:
|
|
|
25-Sep-03
|
Release date:
|
30-Mar-04
|
|
|
|
|
|
|
PROCHECK
|

|
|
|
|
|
Headers
|
|
|
|
References
|
|
|
|
|
|
|
|
|
|
|
|
Enzyme class:
|
|
Chains A, B, C, D:
E.C.1.2.4.4
- 3-methyl-2-oxobutanoate dehydrogenase (2-methylpropanoyl-transferring).
|
|
|
|
|
|
|
Pathway:
|
|
Oxo-acid dehydrogenase complexes
|
|
|
|
|
|
Reaction:
|
|
N6-[(R)-lipoyl]-L-lysyl-[protein] + 3-methyl-2-oxobutanoate + H+ = N6-[(R)-S(8)-2-methylpropanoyldihydrolipoyl]-L-lysyl-[protein] + CO2
|
|
|
|
|
|
N(6)-[(R)-lipoyl]-L-lysyl-[protein]
|
+
|
3-methyl-2-oxobutanoate
Bound ligand (Het Group name = )
matches with 60.00% similarity
|
+
|
H(+)
|
=
|
N(6)-[(R)-S(8)-2-methylpropanoyldihydrolipoyl]-L-lysyl-[protein]
|
+
|
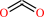 CO2
CO2
|
|
|
|
|
|
|
|
|
|
Cofactor:
|
|
Thiamine diphosphate
|
|
|
|
|
|
Thiamine diphosphate
Bound ligand (Het Group name =
TPP)
corresponds exactly
|
|
|
|
|
|
|
Molecule diagrams generated from .mol files obtained from the
KEGG ftp site
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
| |
|
|
| |
|
DOI no:
|
J Mol Biol
337:1011-1033
(2004)
|
|
PubMed id:
|
|
|
|
|
|
| |
|
Ligand-induced conformational changes and a reaction intermediate in branched-chain 2-oxo acid dehydrogenase (E1) from Thermus thermophilus HB8, as revealed by X-ray crystallography.
|
|
T.Nakai,
N.Nakagawa,
N.Maoka,
R.Masui,
S.Kuramitsu,
N.Kamiya.
|
|
|
|
|
| |
ABSTRACT
|
|
|
|
| |
|
|
The alpha(2)beta(2) tetrameric E1 component of the branched-chain 2-oxo acid
(BCOA) dehydrogenase multienzyme complex is a thiamin diphosphate
(ThDP)-dependent enzyme. E1 catalyzes the decarboxylation of a BCOA concomitant
with the formation of the alpha-carbanion/enamine intermediate,
2-(1-hydroxyalkyl)-ThDP, followed by transfer of the 1-hydroxyalkyl group to the
distal sulfur atom on the lipoamide of the E2 component. In order to elucidate
the catalytic mechanism of E1, the alpha- and beta-subunits of E1 from Thermus
thermophilus HB8 have been co-expressed in Escherichia coli, purified and
crystallized as a stable complex, and the following crystal structures have been
analyzed: the apoenzyme (E1(apo)), the holoenzyme (E1(holo)), E1(holo) in
complex with the substrate analogue 4-methylpentanoate (MPA) as an ES complex
model, and E1(holo) in complex with 4-methyl-2-oxopentanoate (MOPA) as the
alpha-carbanion/enamine intermediate (E1(ceim)). Binding of cofactors to E1(apo)
induces a disorder-order transition in two loops adjacent to the active site.
Furthermore, upon binding of MPA to E1(holo), the loop comprised of
Gly121beta-Gln131beta moves close to the active site and interacts with MPA. The
carboxylate group of MPA is recognized mainly by Tyr86beta and N4' of ThDP. The
hydrophobic moiety of MPA is recognized by Phe66alpha, Tyr95alpha, Met128alpha
and His131alpha. As an intermediate, MOPA is decarboxylated and covalently
linked to ThDP, and the conformation of the protein loop is almost the same as
in the substrate-free (holoenzyme) form. These results suggest that E1 undergoes
an open-closed conformational change upon formation of the ES complex with a
BCOA, and the mobile region participates in the recognition of the carboxylate
group of the BCOA. ES complex models of E1(holo).MOPA and of E1(ceim).lipoamide
built from the above structures suggest that His273alpha and His129beta' are
potential proton donors to the carbonyl group of a BCOA and to the proximal
sulfur atom on the lipoamide, respectively.
|
|
|
|
|
|
| |
Selected figure(s)
|
|
|
|
| |
|
|

|
|

|
|
Figure 1.
Figure 1. Stereoview of the Tth E1 structure. A, Overall
structure of the a[2]b[2] tetramer in the a-carbanion/enamine
intermediate; B, a-subunit; C, b-subunit. The orientations in B
and C are the same as that in A. Ligand molecules are
represented by spheres (orange for Mg2+, yellow for ThDP and
green for HMB group) and those in B and C are shown as
semi-transparent to clarify inner structures. Disordered regions
in E1[apo], which include the STSH motif (see the text and
Figure 3D), are shown in magenta. The substrate-binding loop
(see the text and Figure 6) that undergoes conformational change
upon binding of MPA is shown in cyan. The designation of
secondary structures of Tth E1 is according to those of Ppu E1,
resulting in the absence of helix 1.
|
|
Figure 6.
Figure 6. Conformational changes upon binding of the
substrate analogue MPA. Stereoview of the structure around the
substrate-binding loop for E1[holo] (A), E1[holo]·MPA (B)
and E1[ceim] (C). Carbon atoms of each subunit are colored
according to Figure 1A, whereas a and b in residue labels are
omitted. Water molecules are shown as red spheres. Broken lines
represent hydrogen bonds.
|
|
|
|
|
|
| |
The above figures are
reprinted
by permission from Elsevier:
J Mol Biol
(2004,
337,
1011-1033)
copyright 2004.
|
|
| |
Figures were
selected
by the author.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
 |
 |
|
Literature references that cite this PDB file's key reference
|
|
|
| |
PubMed id
|
|
Reference
|
|
|
|
|
|
X.Y.Pei,
K.M.Erixon,
B.F.Luisi,
and
F.J.Leeper
(2010).
Structural insights into the prereaction state of pyruvate decarboxylase from Zymomonas mobilis .
|
| |
Biochemistry,
49,
1727-1736.
|
|
|
PDB codes:
|
|
|
|
|
|
|
|
S.Kale,
and
F.Jordan
(2009).
Conformational ensemble modulates cooperativity in the rate-determining catalytic step in the E1 component of the Escherichia coli pyruvate dehydrogenase multienzyme complex.
|
| |
J Biol Chem,
284,
33122-33129.
|
|
|
|
|
|
|
M.Kato,
R.M.Wynn,
J.L.Chuang,
S.C.Tso,
M.Machius,
J.Li,
and
D.T.Chuang
(2008).
Structural basis for inactivation of the human pyruvate dehydrogenase complex by phosphorylation: role of disordered phosphorylation loops.
|
| |
Structure,
16,
1849-1859.
|
|
|
PDB codes:
|
|
|
|
|
|
|
|
T.Nakai,
S.Kuramitsu,
and
N.Kamiya
(2008).
Structural bases for the specific interactions between the E2 and E3 components of the Thermus thermophilus 2-oxo acid dehydrogenase complexes.
|
| |
J Biochem,
143,
747-758.
|
|
|
|
|
|
|
V.I.Bunik,
and
D.Degtyarev
(2008).
Structure-function relationships in the 2-oxo acid dehydrogenase family: substrate-specific signatures and functional predictions for the 2-oxoglutarate dehydrogenase-like proteins.
|
| |
Proteins,
71,
874-890.
|
|
|
|
|
|
|
X.Y.Pei,
C.M.Titman,
R.A.Frank,
F.J.Leeper,
and
B.F.Luisi
(2008).
Snapshots of catalysis in the E1 subunit of the pyruvate dehydrogenase multienzyme complex.
|
| |
Structure,
16,
1860-1872.
|
|
|
PDB codes:
|
|
|
|
|
|
|
|
N.Nemeria,
S.Chakraborty,
A.Baykal,
L.G.Korotchkina,
M.S.Patel,
and
F.Jordan
(2007).
The 1',4'-iminopyrimidine tautomer of thiamin diphosphate is poised for catalysis in asymmetric active centers on enzymes.
|
| |
Proc Natl Acad Sci U S A,
104,
78-82.
|
|
|
|
|
|
|
K.Chandrasekhar,
P.Arjunan,
M.Sax,
N.Nemeria,
F.Jordan,
and
W.Furey
(2006).
Active-site changes in the pyruvate dehydrogenase multienzyme complex E1 apoenzyme component from Escherichia coli observed at 2.32 A resolution.
|
| |
Acta Crystallogr D Biol Crystallogr,
62,
1382-1386.
|
|
|
PDB code:
|
|
|
|
|
|
|
|
P.Arjunan,
M.Sax,
A.Brunskill,
K.Chandrasekhar,
N.Nemeria,
S.Zhang,
F.Jordan,
and
W.Furey
(2006).
A thiamin-bound, pre-decarboxylation reaction intermediate analogue in the pyruvate dehydrogenase E1 subunit induces large scale disorder-to-order transformations in the enzyme and reveals novel structural features in the covalently bound adduct.
|
| |
J Biol Chem,
281,
15296-15303.
|
|
|
PDB codes:
|
|
|
|
|
|
|
|
C.L.Berthold,
P.Moussatche,
N.G.Richards,
and
Y.Lindqvist
(2005).
Structural basis for activation of the thiamin diphosphate-dependent enzyme oxalyl-CoA decarboxylase by adenosine diphosphate.
|
| |
J Biol Chem,
280,
41645-41654.
|
|
|
PDB code:
|
|
|
|
|
|
|
|
M.E.Schreiner,
D.Fiur,
J.Holátko,
M.Pátek,
and
B.J.Eikmanns
(2005).
E1 enzyme of the pyruvate dehydrogenase complex in Corynebacterium glutamicum: molecular analysis of the gene and phylogenetic aspects.
|
| |
J Bacteriol,
187,
6005-6018.
|
|
|
|
|
|
|
N.Nemeria,
K.Tittmann,
E.Joseph,
L.Zhou,
M.B.Vazquez-Coll,
P.Arjunan,
G.Hübner,
W.Furey,
and
F.Jordan
(2005).
Glutamate 636 of the Escherichia coli pyruvate dehydrogenase-E1 participates in active center communication and behaves as an engineered acetolactate synthase with unusual stereoselectivity.
|
| |
J Biol Chem,
280,
21473-21482.
|
|
|
|
|
|
|
R.A.Frank,
J.V.Pratap,
X.Y.Pei,
R.N.Perham,
and
B.F.Luisi
(2005).
The molecular origins of specificity in the assembly of a multienzyme complex.
|
| |
Structure,
13,
1119-1130.
|
|
|
PDB code:
|
|
|
|
|
|
|
|
R.A.Frank,
C.M.Titman,
J.V.Pratap,
B.F.Luisi,
and
R.N.Perham
(2004).
A molecular switch and proton wire synchronize the active sites in thiamine enzymes.
|
| |
Science,
306,
872-876.
|
|
|
PDB codes:
|
|
|
|
|
|
|
|
R.M.Wynn,
M.Kato,
M.Machius,
J.L.Chuang,
J.Li,
D.R.Tomchick,
and
D.T.Chuang
(2004).
Molecular mechanism for regulation of the human mitochondrial branched-chain alpha-ketoacid dehydrogenase complex by phosphorylation.
|
| |
Structure,
12,
2185-2196.
|
|
|
PDB codes:
|
|
|
|
|
|
|
The most recent references are shown first.
Citation data come partly from CiteXplore and partly
from an automated harvesting procedure. Note that this is likely to be
only a partial list as not all journals are covered by
either method. However, we are continually building up the citation data
so more and more references will be included with time.
Where a reference describes a PDB structure, the PDB
codes are
shown on the right.
|
 |



 Links
Links













