 |
PDBsum entry 4hi3
 |
|
|
|
|
|
|
Enzyme class 1:
|
|
E.C.2.7.7.50
- mRNA guanylyltransferase.
|
|
|
|
|
|
|
Reaction:
|
|
a 5'-end diphospho-ribonucleoside in mRNA + GTP + H+ = a 5'-end (5'-triphosphoguanosine)-ribonucleoside in mRNA + diphosphate
|
|
|
|
|
|
5'-end diphospho-ribonucleoside in mRNA
|
+
|
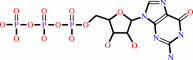 GTP
GTP
|
+
|
H(+)
|
=
|
5'-end (5'-triphosphoguanosine)-ribonucleoside in mRNA
|
+
|
 diphosphate
diphosphate
|
|
|
|
|
|
|
|
|
|
Enzyme class 2:
|
|
E.C.3.4.19.12
- ubiquitinyl hydrolase 1.
|
|
|
|
|
|
|
Reaction:
|
|
Thiol-dependent hydrolysis of ester, thiolester, amide, peptide and isopeptide bonds formed by the C-terminal Gly of ubiquitin (a 76-residue protein attached to proteins as an intracellular targeting signal).
|
|
|
|
|
|
Enzyme class 3:
|
|
E.C.3.4.22.-
- ?????
|
|
|
|
|
|
|
Enzyme class 4:
|
|
E.C.3.4.22.69
- Sars coronavirus main proteinase.
|
|
|
|
|
|
|
|
|
|
Note, where more than one E.C. class is given (as above), each may
correspond to a different protein domain or, in the case of polyprotein
precursors, to a different mature protein.
|
|
|
|
Molecule diagrams generated from .mol files obtained from the
KEGG ftp site
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
| |
|
|
| |
|
DOI no:
|
Acta Crystallogr D Biol Crystallogr
69:747-755
(2013)
|
|
PubMed id:
|
|
|
|
|
|
| |
|
Mechanism for controlling the monomer-dimer conversion of SARS coronavirus main protease.
|
|
C.G.Wu,
S.C.Cheng,
S.C.Chen,
J.Y.Li,
Y.H.Fang,
Y.H.Chen,
C.Y.Chou.
|
|
|
|
|
| |
ABSTRACT
|
|
|
|
| |
|
|
The Severe acute respiratory syndrome coronavirus (SARS-CoV) main protease
(M(pro)) cleaves two virion polyproteins (pp1a and pp1ab); this essential
process represents an attractive target for the development of anti-SARS drugs.
The functional unit of M(pro) is a homodimer and each subunit contains a
His41/Cys145 catalytic dyad. Large amounts of biochemical and structural
information are available on M(pro); nevertheless, the mechanism by which
monomeric M(pro) is converted into a dimer during maturation still remains
poorly understood. Previous studies have suggested that a C-terminal residue,
Arg298, interacts with Ser123 of the other monomer in the dimer, and mutation of
Arg298 results in a monomeric structure with a collapsed substrate-binding
pocket. Interestingly, the R298A mutant of M(pro) shows a reversible
substrate-induced dimerization that is essential for catalysis. Here, the
conformational change that occurs during substrate-induced dimerization is
delineated by X-ray crystallography. A dimer with a mutual orientation of the
monomers that differs from that of the wild-type protease is present in the
asymmetric unit. The presence of a complete substrate-binding pocket and
oxyanion hole in both protomers suggests that they are both catalytically
active, while the two domain IIIs show minor reorganization. This structural
information offers valuable insights into the molecular mechanism associated
with substrate-induced dimerization and has important implications with respect
to the maturation of the enzyme.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
 |
 |
|


 Links
Links



























