 |
PDBsum entry 3eyc

|
|
|
|
|
|

|
|

|
|

|
|

|
|

|
|

|
|

|
|

|
|

|
|
|
 |

|

|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Transport protein
|
PDB id
|
|
|
|
3eyc
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|

|

|

|
|
|
|
|
|
References listed in PDB file
|
|
|
Key reference
|
|
|
Title
|
|
A new crystal form of human tear lipocalin reveals high flexibility in the loop region and induced fit in the ligand cavity.
|
|
|
Authors
|
|
D.A.Breustedt,
L.Chatwell,
A.Skerra.
|
|
|
Ref.
|
|
Acta Crystallogr D Biol Crystallogr, 2009,
65,
1118-1125.
[DOI no: ]
|
|
|
PubMed id
|
|
|
|
|
|
Abstract
|
|
|
Tear lipocalin (TLC) with the bound artificial ligand 1,4-butanediol has been
crystallized in space group P2(1) with four protein molecules in the asymmetric
unit and its X-ray structure has been solved at 2.6 A resolution. TLC is a
member of the lipocalin family that binds ligands with diverse chemical
structures, such as fatty acids, phospholipids and cholesterol as well as
microbial siderophores and the antibiotic rifampin. Previous X-ray structural
analysis of apo TLC crystallized in space group C2 revealed a rather large
bifurcated ligand pocket and a partially disordered loop region at the entrace
to the cavity. Analysis of the P2(1) crystal form uncovered major conformational
changes (i) in beta-strands B, C and D, (ii) in loops 1, 2 and 4 at the open end
of the beta-barrel and (iii) in the extended C-terminal segment, which is
attached to the beta-barrel via a disulfide bridge. The structural comparison
indicates high conformational plasticity of the loop region as well as of deeper
parts of the ligand pocket, thus allowing adaptation to ligands that differ
vastly in size and shape. This illustrates a mechanism for promiscuity in ligand
recognition which may also be relevant for some other physiologically important
members of the lipocalin protein family.
|
|
|
|

|
|
Figure 3.
Figure 3 Comparison of the TLC structures obtained from two
different crystal forms. Molecules were superimposed using the
C^  positions
of the 58 conserved positions
of the 58 conserved 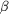 -barrel
residues (as in Fig. 1-b). (a) Superposition of TLC crystallized
in space group P2[1] (chain A, blue) and in space group C2 (PDB
code 1xki ; gold, with modelled residues in grey; Breustedt et
al., 2005[Breustedt, D. A., Korndörfer, I. P., Redl, B. &
Skerra, A. (2005). J. Biol. Chem. 280, 484-493.]). The bound
1,4-butanediol and the hydrogen-bonded water molecule are
represented as spheres, while the conserved disulfide bond is
shown in stick representation (white). Most of the -strands
are elongated towards the open end of the -barrel
in the P2[1] structure. (b) Pairwise C^ -atom
distances between the two different TLC crystal structures after
superposition of the 58 conserved C^ positions
of the -barrel,
resulting in an overall C^ r.m.s.d.
value of 1.86 Å ( -strands
A-H are labelled as bars; loops 1, 2, 3 and 4 connect strands A
and B, C and D, E and F, and G and H, respectively). The largest
deviations between the two crystal structures occur in -strands
B, C and D and in the neighbouring loops 1 and 2 at the open end
of the -barrel.
(c) The electrostatic interactions that may trigger the two
alternative conformations of loop 1. In the P2[1] structure
(dark grey) residues Glu27 and Lys108 (green) at the tips of the
adjacent loops 1 and 4 make an electrostatic contact that fixes
the `open' conformation of loop 1 (blue). In contrast, the
`closed' conformation of this loop in the C2 structure (light
grey; loop 1 coloured gold, modelled residues coloured yellow)
is stabilized by a similar interaction between residues Glu34
and Lys114 (orange). (d) Flexibility of the region around the
disulfide bridge. While the disulfide bond between Cys61 and
Cys153 is well ordered in the P2[1] structure (dark grey; loop 2
and the C-terminal peptide segment coloured blue, side chains
coloured violet), only residue Cys61 was resolved in the C2
structure (light grey; loop 2 and the C-terminal peptide segment
coloured gold, modelled residues coloured yellow, Cys side chain
coloured orange) and appears shifted outward by 5.6 Å
compared with the P2[1] structure (red dotted line between the
corresponding C^ positions).
Nevertheless, the C^ distance
between Cys61 in the C2 structure and Cys153 in the superimposed
P2[1] structure is only 0.3 Å larger than the distance
between Cys61 and Cys153 within the P2[1] structure (green
dotted lines). By assuming a more extended backbone conformation
at residue Glu151 (side chain displayed in blue), the
(unresolved) C-terminal peptide segment of the C2 structure
could easily span the 0.3 Å distance and move Cys153
sufficiently close to Cys61, indicating that the disulfide bond
is probably also formed in the C2 crystal (despite the differing
conformation of loop 2) but is not visible in the electron
density owing to local structural disorder. -barrel
residues (as in Fig. 1-b). (a) Superposition of TLC crystallized
in space group P2[1] (chain A, blue) and in space group C2 (PDB
code 1xki ; gold, with modelled residues in grey; Breustedt et
al., 2005[Breustedt, D. A., Korndörfer, I. P., Redl, B. &
Skerra, A. (2005). J. Biol. Chem. 280, 484-493.]). The bound
1,4-butanediol and the hydrogen-bonded water molecule are
represented as spheres, while the conserved disulfide bond is
shown in stick representation (white). Most of the -strands
are elongated towards the open end of the -barrel
in the P2[1] structure. (b) Pairwise C^ -atom
distances between the two different TLC crystal structures after
superposition of the 58 conserved C^ positions
of the -barrel,
resulting in an overall C^ r.m.s.d.
value of 1.86 Å ( -strands
A-H are labelled as bars; loops 1, 2, 3 and 4 connect strands A
and B, C and D, E and F, and G and H, respectively). The largest
deviations between the two crystal structures occur in -strands
B, C and D and in the neighbouring loops 1 and 2 at the open end
of the -barrel.
(c) The electrostatic interactions that may trigger the two
alternative conformations of loop 1. In the P2[1] structure
(dark grey) residues Glu27 and Lys108 (green) at the tips of the
adjacent loops 1 and 4 make an electrostatic contact that fixes
the `open' conformation of loop 1 (blue). In contrast, the
`closed' conformation of this loop in the C2 structure (light
grey; loop 1 coloured gold, modelled residues coloured yellow)
is stabilized by a similar interaction between residues Glu34
and Lys114 (orange). (d) Flexibility of the region around the
disulfide bridge. While the disulfide bond between Cys61 and
Cys153 is well ordered in the P2[1] structure (dark grey; loop 2
and the C-terminal peptide segment coloured blue, side chains
coloured violet), only residue Cys61 was resolved in the C2
structure (light grey; loop 2 and the C-terminal peptide segment
coloured gold, modelled residues coloured yellow, Cys side chain
coloured orange) and appears shifted outward by 5.6 Å
compared with the P2[1] structure (red dotted line between the
corresponding C^ positions).
Nevertheless, the C^ distance
between Cys61 in the C2 structure and Cys153 in the superimposed
P2[1] structure is only 0.3 Å larger than the distance
between Cys61 and Cys153 within the P2[1] structure (green
dotted lines). By assuming a more extended backbone conformation
at residue Glu151 (side chain displayed in blue), the
(unresolved) C-terminal peptide segment of the C2 structure
could easily span the 0.3 Å distance and move Cys153
sufficiently close to Cys61, indicating that the disulfide bond
is probably also formed in the C2 crystal (despite the differing
conformation of loop 2) but is not visible in the electron
density owing to local structural disorder.
|
|
|
|
|
|
The above figure is
reprinted
by permission from the IUCr:
Acta Crystallogr D Biol Crystallogr
(2009,
65,
1118-1125)
copyright 2009.
|
|
|
|
|
|
|


