 |
PDBsum entry 1j6y

|
|
|
|
|
 |
Contents |
 |
|
|
|
|
|
|
|
|
* Residue conservation analysis
|
|
|
|
|
|
|
|
|
|
|
Enzyme class:
|
|
E.C.5.2.1.8
- peptidylprolyl isomerase.
|
|
|
|
|
|
|
Reaction:
|
|
[protein]-peptidylproline (omega=180) = [protein]-peptidylproline (omega=0)
|
|
|
|
|
|
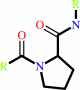 Peptidylproline (omega=180)
Peptidylproline (omega=180)
|
=
|
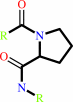 peptidylproline (omega=0)
peptidylproline (omega=0)
|
|
|
|
|
|
|
|
|
|
|
|
|
Molecule diagrams generated from .mol files obtained from the
KEGG ftp site
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
| |
|
|
| |
|
DOI no:
|
J Mol Biol
320:321-332
(2002)
|
|
PubMed id:
|
|
|
|
|
|
| |
|
Solution structure of the single-domain prolyl cis/trans isomerase PIN1At from Arabidopsis thaliana.
|
|
I.Landrieu,
J.M.Wieruszeski,
R.Wintjens,
D.Inzé,
G.Lippens.
|
|
|
|
|
| |
ABSTRACT
|
|
|
|
| |
|
|
The 119-amino acid residue prolyl cis/trans isomerase from Arabidopsis thaliana
(PIN1At) is similar to the catalytic domain of the human hPIN1. However, PIN1At
lacks the N-terminal WW domain that appears to be essential for the hPIN1
function. Here, the solution structure of PIN1At was determined by
three-dimensional nuclear magnetic resonance spectroscopy. The PIN1At fold could
be superimposed on that of the catalytic domain of hPIN1 and had a 19 residue
flexible loop located between strand beta1 and helix alpha1. The dynamical
features of this beta1/alpha1-loop, which are characteristic for a region
involved in protein-protein interactions, led to exchange broadening in the NMR
spectra. When sodium sulfate salt was added to the protein sample, the
beta1/alpha1 loop was stabilized and, hence, a complete backbone resonance
assignment was obtained. Previously, with a phospho-Cdc25 peptide as substrate,
PIN1At had been shown to catalyze the phosphoserine/phosphothreonine prolyl
cis/trans isomerization specifically. To map the catalytic site of PIN1At, the
phospho-Cdc25 peptide or sodium sulfate salt was added in excess to the protein
and chemical shift changes in the backbone amide protons were monitored in the
(1)H(N)-(15)N heteronuclear single quantum coherence spectrum. The peptide
caused perturbations in the loops between helix alpha4 and strand beta3, between
strands beta3 and beta4, in the alpha3 helix, and in the beta1/alpha1 loop. The
amide groups of the residues Arg21 and Arg22 showed large chemical shift
perturbations upon phospho-Cdc25 peptide or sulfate addition. We conclude that
this basic cluster formed by Arg21 and Arg22, both located in the beta1/alpha1
loop, is homologous to that found in the hPIN1 crystal structure (Arg68 and
Arg69), which also is involved in sulfate ion binding. We showed that the
sulfate group competed for the interaction between PIN1At and the phospho-Cdc25
peptide. In the absence of the WW domain, three hydrophobic residues (Ile33,
Ile34, and Leu35) located in the long flexible loop and specific for the plant
PIN-type peptidyl prolyl cis/trans isomerases (PPIases) could be an additional
interaction site in PIN1At. However, phospho-peptide addition did not affect the
resonances of these residues significantly. Electrostatic potential calculations
revealed a negatively charged area not found in hPIN1 on the PIN1At molecular
surface, which corresponds to the surface shielded by the WW domain in hPIN1.
Based on our experimental results and the molecular specificities of the PIN1At
enzyme, functional implications of the lack of WW domains in this plant PIN-type
PPIase will be discussed.
|
|
|
|
|
|
| |
Selected figure(s)
|
|
|
|
| |
|
|

|
|

|
|
Figure 3.
Figure 3. Ribbon representations of (a) hPIN1 (residues
9-37 and 50-163) and (b) representative NMR PIN1At conformer
(residues 6-120). The coordinates of hPIN1 are from PDB:1PIN.5
Secondary structure elements are colored and labeled. The
sulfate molecule and the Ala-Pro dipeptide are represented by
the space-filling model and are colored blue and green,
respectively. The picture in (a) was obtained using a
combination of MOLSCRIPT and Raster3D programs.
|
|
Figure 6.
Figure 6. Accessible molecular surface representation of
(a) hPIN1 and (b) PIN1At in similar orientation, colored
according to the electrostatic potential and displayed with
GRASP program. Color codes for the electric potential are -5kT/e
(red, acidic residues), 0kT/e (white), +5kT/e (blue, basic
residues). The WW domain of hPIN1 (a) is drawn as a yellow line.
The view is along the surface shielded in hPIN1 by the WW domain
and corresponds to a 20° rotation to the left around the
ordinate when compared with Figure 3.
|
|
|
|
|
|
| |
The above figures are
reprinted
by permission from Elsevier:
J Mol Biol
(2002,
320,
321-332)
copyright 2002.
|
|
| |
Figures were
selected
by the author.
|
|
|
|
| |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| |
|
|
|
|
| |
Since this first report of a plant Pin1 homolog without WW domain, other plant parvulins have been reported (see Yao JL, Kops O, Lu PJ, Lu KP.
Functional conservation of phosphorylation-specific prolyl isomerases in
plants.
.)
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
 |
 |
 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Literature references that cite this PDB file's key reference
|
|
|
| |
PubMed id
|
|
Reference
|
|
|
|
|
|
O.Heikkinen,
R.Seppala,
H.Tossavainen,
S.Heikkinen,
H.Koskela,
P.Permi,
and
I.Kilpeläinen
(2009).
Solution structure of the parvulin-type PPIase domain of Staphylococcus aureus PrsA--implications for the catalytic mechanism of parvulins.
|
| |
BMC Struct Biol,
9,
17.
|
|
|
PDB code:
|
|
|
|
|
|
|
|
J.W.Mueller,
and
P.Bayer
(2008).
Small family with key contacts: par14 and par17 parvulin proteins, relatives of pin1, now emerge in biomedical research.
|
| |
Perspect Medicin Chem,
2,
11-20.
|
|
|
|
|
|
|
G.Lippens,
I.Landrieu,
and
C.Smet
(2007).
Molecular mechanisms of the phospho-dependent prolyl cis/trans isomerase Pin1.
|
| |
FEBS J,
274,
5211-5222.
|
|
|
|
|
|
|
A.Kühlewein,
G.Voll,
B.Hernandez Alvarez,
H.Kessler,
G.Fischer,
J.U.Rahfeld,
and
G.Gemmecker
(2004).
Solution structure of Escherichia coli Par10: The prototypic member of the Parvulin family of peptidyl-prolyl cis/trans isomerases.
|
| |
Protein Sci,
13,
2378-2387.
|
|
|
PDB codes:
|
|
|
|
|
|
|
|
H.Mira,
M.Vilar,
V.Esteve,
M.Martinell,
M.J.Kogan,
E.Giralt,
D.Salom,
I.Mingarro,
L.Peñarrubia,
and
E.Pérez-Payá
(2004).
Ionic self-complementarity induces amyloid-like fibril formation in an isolated domain of a plant copper metallochaperone protein.
|
| |
BMC Struct Biol,
4,
7.
|
|
|
|
|
|
|
D.M.Jacobs,
K.Saxena,
M.Vogtherr,
P.Bernado,
M.Pons,
and
K.M.Fiebig
(2003).
Peptide binding induces large scale changes in inter-domain mobility in human Pin1.
|
| |
J Biol Chem,
278,
26174-26182.
|
|
|
|
|
|
|
E.Bayer,
S.Goettsch,
J.W.Mueller,
B.Griewel,
E.Guiberman,
L.M.Mayr,
and
P.Bayer
(2003).
Structural analysis of the mitotic regulator hPin1 in solution: insights into domain architecture and substrate binding.
|
| |
J Biol Chem,
278,
26183-26193.
|
|
|
PDB codes:
|
|
|
|
|
|
|
The most recent references are shown first.
Citation data come partly from CiteXplore and partly
from an automated harvesting procedure. Note that this is likely to be
only a partial list as not all journals are covered by
either method. However, we are continually building up the citation data
so more and more references will be included with time.
Where a reference describes a PDB structure, the PDB
code is
shown on the right.
|
|


 Links
Links










