 |
PDBsum entry 4ccn

|
|
|
|
|
|

|

|

|

|
|

|
|

|
|

|
|

|
|

|
|

|
|

|
|
|
 |

|

|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Oxidoreductase
|
PDB id
|
|
|
|
4ccn
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|

|

|

|
|
|
|
|
|
|
PDB id:
|
|
|
|
| Name: |
|
Oxidoreductase
|
|
|
Title:
|
|
60s ribosomal protein l8 histidine hydroxylase (no66 l299c/c300s) in complex with mn(ii), n-oxalylglycine (nog) and 60s ribosomal protein l8 (rpl8 g220c) peptide fragment (complex-2)
|
|
Structure:
|
|
Bifunctional lysine-specific demethylase and histidyl- hydroxylase no66. Chain: a, b. Fragment: catalytic domain, residues 183-641. Synonym: 60s ribosomal protein l8 histidine hydroxylase, histone lysine demethylase no66, myc-associated protein with jmjc domain, nucleolar protein 66, hsno66, ribosomal oxygenase no66, rox. Engineered: yes. 60s ribosomal protein l8.
|
|
Source:
|
|
Homo sapiens. Human. Organism_taxid: 9606. Expressed in: escherichia coli. Expression_system_taxid: 469008. Synthetic: yes. Organism_taxid: 9606
|
|
Resolution:
|
|
|
2.23Å
|
R-factor:
|
0.238
|
R-free:
|
0.248
|
|
|
Authors:
|
|
R.Chowdhury,C.J.Schofield
|
|
Key ref:
|
|
R.Chowdhury
et al.
(2014).
Ribosomal oxygenases are structurally conserved from prokaryotes to humans.
Nature,
510,
422-426.
PubMed id:
DOI:
|
|
|
Date:
|
|
|
23-Oct-13
|
Release date:
|
14-May-14
|
|
|
|
|
|
|
PROCHECK
|

|
|
|
|
|
Headers
|
|
|
|
References
|
|
|
|
|
|

|
|
Q9H6W3
(RIOX1_HUMAN) -
Ribosomal oxygenase 1 from Homo sapiens
|
|
|
|
Seq:
Struc:
|
|
|
|
641 a.a.
458 a.a.*
|
|

|

|
|
|
|
|
|
|
|
|
|
|
|
Key: |
 |
PfamA domain |
|
|
 |
Secondary structure |
|
 |
CATH domain |
|
|
*
PDB and UniProt seqs differ
at 4 residue positions (black
crosses)
|
|
|
|
|
|
|
|
|
|
|
|
|
Enzyme class 2:
|
|
E.C.1.14.11.27
- [histone H3]-dimethyl-L-lysine(36) demethylase.
|
|
|
|
|
|
|
Reaction:
|
|
N6,N6-dimethyl-L-lysyl36-[histone H3] + 2 2-oxoglutarate + 2 O2 = L-lysyl36-[histone H3] + 2 formaldehyde + 2 succinate + 2 CO2
|
|
|
|
|
|
N(6),N(6)-dimethyl-L-lysyl(36)-[histone H3]
|
+
|
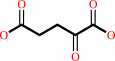 2
×
2-oxoglutarate
2
×
2-oxoglutarate
|
+
|
 2
×
O2
2
×
O2
|
=
|
L-lysyl(36)-[histone H3]
|
+
|
 2
×
formaldehyde
2
×
formaldehyde
|
+
|
 2
×
succinate
2
×
succinate
|
+
|
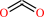 2
×
CO2
2
×
CO2
|
|
|
|
|
|
|
|
|
|
Cofactor:
|
|
Fe(2+)
|
|
|
|
|
|
Enzyme class 3:
|
|
E.C.1.14.11.79
- protein-L-histidine (3S)-3-hydroxylase.
|
|
|
|
|
|
|
Reaction:
|
|
L-histidyl-[protein] + 2-oxoglutarate + O2 = (3S)-3-hydroxy-L-histidyl- [protein] + succinate + CO2
|
|
|
|
|
|
L-histidyl-[protein]
Bound ligand (Het Group name = )
matches with 58.33% similarity
|
+
|
2
×
2-oxoglutarate
|
+
|
2
×
O2
|
=
|
(3S)-3-hydroxy-L-histidyl- [protein]
|
+
|
2
×
succinate
Bound ligand (Het Group name = )
matches with 50.00% similarity
|
+
|
2
×
CO2
|
|
|
|
|
|
|
|
|
|
|
|
|
Note, where more than one E.C. class is given (as above), each may
correspond to a different protein domain or, in the case of polyprotein
precursors, to a different mature protein.
|
|
|
|
Molecule diagrams generated from .mol files obtained from the
KEGG ftp site
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
| |
|
|
| |
|
DOI no:
|
Nature
510:422-426
(2014)
|
|
PubMed id:
|
|
|
|
|
|
| |
|
Ribosomal oxygenases are structurally conserved from prokaryotes to humans.
|
|
R.Chowdhury,
R.Sekirnik,
N.C.Brissett,
T.Krojer,
C.H.Ho,
S.S.Ng,
I.J.Clifton,
W.Ge,
N.J.Kershaw,
G.C.Fox,
J.R.Muniz,
M.Vollmar,
C.Phillips,
E.S.Pilka,
K.L.Kavanagh,
F.von Delft,
U.Oppermann,
M.A.McDonough,
A.J.Doherty,
C.J.Schofield.
|
|
|
|
|
| |
ABSTRACT
|
|
|
|
| |
|
|
2-Oxoglutarate (2OG)-dependent oxygenases have important roles in the regulation
of gene expression via demethylation of N-methylated chromatin components and in
the hydroxylation of transcription factors and splicing factor proteins.
Recently, 2OG-dependent oxygenases that catalyse hydroxylation of transfer RNA
and ribosomal proteins have been shown to be important in translation relating
to cellular growth, TH17-cell differentiation and translational accuracy. The
finding that ribosomal oxygenases (ROXs) occur in organisms ranging from
prokaryotes to humans raises questions as to their structural and evolutionary
relationships. In Escherichia coli, YcfD catalyses arginine hydroxylation in the
ribosomal protein L16; in humans, MYC-induced nuclear antigen (MINA53; also
known as MINA) and nucleolar protein 66 (NO66) catalyse histidine hydroxylation
in the ribosomal proteins RPL27A and RPL8, respectively. The functional
assignments of ROXs open therapeutic possibilities via either ROX inhibition or
targeting of differentially modified ribosomes. Despite differences in the
residue and protein selectivities of prokaryotic and eukaryotic ROXs, comparison
of the crystal structures of E. coli YcfD and Rhodothermus marinus YcfD with
those of human MINA53 and NO66 reveals highly conserved folds and novel
dimerization modes defining a new structural subfamily of 2OG-dependent
oxygenases. ROX structures with and without their substrates support their
functional assignments as hydroxylases but not demethylases, and reveal how the
subfamily has evolved to catalyse the hydroxylation of different residue side
chains of ribosomal proteins. Comparison of ROX crystal structures with those of
other JmjC-domain-containing hydroxylases, including the hypoxia-inducible
factor asparaginyl hydroxylase FIH and histone N(ε)-methyl lysine demethylases,
identifies branch points in 2OG-dependent oxygenase evolution and distinguishes
between JmjC-containing hydroxylases and demethylases catalysing modifications
of translational and transcriptional machinery. The structures reveal that new
protein hydroxylation activities can evolve by changing the coordination
position from which the iron-bound substrate-oxidizing species reacts. This
coordination flexibility has probably contributed to the evolution of the wide
range of reactions catalysed by oxygenases.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
 |
 |
|



 Links
Links






