
|
|
|
|
|
 |
Contents |
 |
|
|
|
|
|
|
|
|
|
|
|





 (+ 2 more)
478 a.a.
(+ 2 more)
478 a.a.
|
|
|
|
|
|
|
|
|
|
|





 (+ 2 more)
410 a.a.
(+ 2 more)
410 a.a.
|
|
|
|
|
|
|
|
|
|
|





 (+ 2 more)
91 a.a.
(+ 2 more)
91 a.a.
|
|
|
|
|
|
|
|
|
|
|
|
|
* Residue conservation analysis
|
|
|
|
|
|
PDB id:
|
|
|
|
| Name: |
|
Ligase
|
|
|
Title:
|
|
Structure of tRNA-dependent amidotransferase gatcab from aquifex aeolicus
|
|
Structure:
|
|
Glutamyl-tRNA(gln) amidotransferase subunit a. Chain: a, d, g, j, m, p, s, v. Synonym: glu-adt subunit a. Engineered: yes. Aspartyl/glutamyl-tRNA(asn/gln) amidotransferase subunit b. Chain: b, e, h, k, n, q, t, w. Engineered: yes. Glutamyl-tRNA(gln) amidotransferase subunit c. Chain: c, f, i, l, o, r, u, x.
|
|
Source:
|
|
Aquifex aeolicus. Organism_taxid: 63363. Strain: vf5. Gene: aq_247, gata, gatca. Expressed in: escherichia coli. Expression_system_taxid: 562. Gene: aq_461, gatb. Gene: aq_2149, gatc.
|
|
Resolution:
|
|
|
3.00Å
|
R-factor:
|
0.265
|
R-free:
|
0.306
|
|
|
Authors:
|
|
J.Wu,W.Bu,K.Sheppard,M.Kitabatake,D.Soll,J.L.Smith
|
Key ref:

|
|
J.Wu
et al.
(2009).
Insights into tRNA-dependent amidotransferase evolution and catalysis from the structure of the Aquifex aeolicus enzyme.
J Mol Biol,
391,
703-716.
PubMed id:
DOI:

|
|
|
Date:
|
|
|
10-Apr-09
|
Release date:
|
21-Jul-09
|
|
|
|
|
|
|
PROCHECK
|

|
|
|
|
|
Headers
|
|
|
|
References
|
|
|
|
|
|

|
|
O66610
(GATA_AQUAE) -
Glutamyl-tRNA(Gln) amidotransferase subunit A from Aquifex aeolicus (strain VF5)
|
|
|
|
Seq:
Struc:
|
|
|
|
478 a.a.
478 a.a.
|
|

|

|

|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Enzyme class 2:
|
|
Chains A, D, G, J, M, P, S, V:
E.C.6.3.5.7
- glutaminyl-tRNA synthase (glutamine-hydrolyzing).
|
|
|
|
|
|
|
Reaction:
|
|
L-glutamyl-tRNA(Gln) + L-glutamine + ATP + H2O = L-glutaminyl-tRNA(Gln) + L-glutamate + ADP + phosphate + H+
|
|
|
|
|
|
L-glutamyl-tRNA(Gln)
Bound ligand (Het Group name = )
corresponds exactly
|
+
|
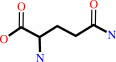 L-glutamine
L-glutamine
|
+
|
ATP
Bound ligand (Het Group name = )
matches with 63.64% similarity
|
+
|
H2O
|
=
|
L-glutaminyl-tRNA(Gln)
Bound ligand (Het Group name = )
corresponds exactly
|
+
|
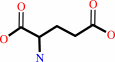 L-glutamate
L-glutamate
|
+
|
 ADP
ADP
|
+
|
 phosphate
phosphate
|
+
|
H(+)
|
|
|
|
|
|
|
|
|
|
Enzyme class 3:
|
|
Chains B, C, E, F, H, I, K, L, N, O, Q, R, T, U, W, X:
E.C.6.3.5.-
- ?????
|
|
|
|
|
|
|
|
|
|
Note, where more than one E.C. class is given (as above), each may
correspond to a different protein domain or, in the case of polyprotein
precursors, to a different mature protein.
|
|
|
|
Molecule diagrams generated from .mol files obtained from the
KEGG ftp site
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
| |
|
|
| |
|
DOI no:
|
J Mol Biol
391:703-716
(2009)
|
|
PubMed id:
|
|
|
|
|
|
| |
|
Insights into tRNA-dependent amidotransferase evolution and catalysis from the structure of the Aquifex aeolicus enzyme.
|
|
J.Wu,
W.Bu,
K.Sheppard,
M.Kitabatake,
S.T.Kwon,
D.Söll,
J.L.Smith.
|
|
|
|
|
| |
ABSTRACT
|
|
|
|
| |
|
|
Many bacteria form Gln-tRNA(Gln) and Asn-tRNA(Asn) by conversion of the
misacylated Glu-tRNA(Gln) and Asp-tRNA(Asn) species catalyzed by the GatCAB
amidotransferase in the presence of ATP and an amide donor (glutamine or
asparagine). Here, we report the crystal structures of GatCAB from the
hyperthermophilic bacterium Aquifex aeolicus, complexed with glutamine,
asparagine, aspartate, ADP, or ATP. In contrast to the Staphylococcus aureus
GatCAB, the A. aeolicus enzyme formed acyl-enzyme intermediates with either
glutamine or asparagine, in line with the equally facile use by the
amidotransferase of these amino acids as amide donors in the transamidation
reaction. A water-filled ammonia channel is open throughout the length of the A.
aeolicus GatCAB from the GatA active site to the synthetase catalytic pocket in
the B-subunit. A non-catalytic Zn(2+) site in the A. aeolicus GatB stabilizes
subunit contacts and the ammonia channel. Judged from sequence conservation in
the known GatCAB sequences, the Zn(2+) binding motif was likely present in the
primordial GatB/E, but became lost in certain lineages (e.g., S. aureus GatB).
Two divalent metal binding sites, one permanent and the other transient, are
present in the catalytic pocket of the A. aeolicus GatB. The two sites enable
GatCAB to first phosphorylate the misacylated tRNA substrate and then amidate
the activated intermediate to form the cognate products, Gln-tRNA(Gln) or
Asn-tRNA(Asn).
|
|
|
|
|
|
| |
Selected figure(s)
|
|
|
|
| |
|
|

|
|

|
|
Figure 3.
Fig. 3. Comparison of amidase active sites. (a) A-subunit of
A. aeolicus GatCAB showing the acyl-enzyme intermediate of
substrate Gln with Ser171. (b) Active site of MAE2 in complex
with product malonate.^23 Arg158 in the amidase core region
interacts with a carboxyl group of malonate (Mal). (c) Active
site of FAAH with the inactivator methoxy arachidonyl
phosphonate (MAP).^26 The phosphonate of the covalent adduct at
nucleophilic Ser241 mimics the tetrahedral intermediate of the
hydrolytic reaction. Aromatic and aliphatic residues in the
substrate binding pocket are indicated. (d) Active site of PAM
in complex with the inhibitor chymostatin (CST).^25 For each
enzyme, the amidase core region (residues 62–192 of GatA,
residues 52–176 of MAE2, residues 132–262 of FAAH, and
residues 113–246 of PAM) is colored blue, and residues outside
the core region are colored green. Residues in the
Ser–cisSer–Lys catalytic scissors of each enzyme and those
interacting with ligands are shown as thin sticks; adducts and
ligands are shown in ball-and-stick form with atomic coloring:
gray, C; red, O; blue, N; orange, P. Hydrogen bonds are shown as
dashed lines.
|
|
Figure 7.
Fig. 7. Proposed reactions at the synthetase active site. (a)
Superposition of synthetase active sites from crystal structures
with informative ligands. The A. aeolicus GatCAB with ATP,
Mn^2+, and Asp (this work) is shown in green with yellow C atoms
for the ligands; the S. aureus GatCAB with ADP and Mg^2+^14 in
orange with orange C for the ligands; and the M.
thermautotrophicus GatDE with tRNA^15 in magenta with yellow C
for tRNA. Mn^2+ ions are shown as purple spheres, Mg^2+ in
orange, and water molecules as red spheres. Substrates ATP, ADP,
Asp, and 3′-CCA of tRNA^Gln are represented as sticks.
Residues interacting with substrates are represented by thick
lines. The ammonia channel (gray surface) enters the synthetase
active site from the right and is continuous with the tRNA
binding site. (b) Model for the activation reaction. ATP is
positioned as in the structure reported here. The terminus of
Asp-tRNA^Asn was modeled based on the GatDE–tRNA complex in
which the 3′-terminal A was disordered. The Asp carboxyl group
is coordinated by the metal in the permanent site, as in the Asp
complex. (c) Model for the amidation complex. The activated
substrate, phosphoryl-Asp-tRNA^Asn, is shifted so that both
phosphate and O^δ coordinate the permanent metal, thereby
positioning the Asp C^γ atom at the exit of the ammonia tunnel,
ready to receive ammonia from the amidase active site. (d)
Schematic diagram of the reaction steps depicted in (b) and (c).
|
|
|
|
|
|
| |
The above figures are
reprinted
by permission from Elsevier:
J Mol Biol
(2009,
391,
703-716)
copyright 2009.
|
|
| |
Figures were
selected
by an automated process.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
 |
 |
|
Literature references that cite this PDB file's key reference
|
|
|
| |
PubMed id
|
|
Reference
|
|
|
|
|
|
A.Nakamura,
K.Sheppard,
J.Yamane,
M.Yao,
D.Söll,
and
I.Tanaka
(2010).
Two distinct regions in Staphylococcus aureus GatCAB guarantee accurate tRNA recognition.
|
| |
Nucleic Acids Res,
38,
672-682.
|
|
|
PDB code:
|
|
|
|
|
|
|
|
M.Blaise,
M.Bailly,
M.Frechin,
M.A.Behrens,
F.Fischer,
C.L.Oliveira,
H.D.Becker,
J.S.Pedersen,
S.Thirup,
and
D.Kern
(2010).
Crystal structure of a transfer-ribonucleoprotein particle that promotes asparagine formation.
|
| |
EMBO J,
29,
3118-3129.
|
|
|
PDB code:
|
|
|
|
|
|
|
|
T.Ito,
and
S.Yokoyama
(2010).
Two enzymes bound to one transfer RNA assume alternative conformations for consecutive reactions.
|
| |
Nature,
467,
612-616.
|
|
|
PDB codes:
|
|
|
|
|
|
|
The most recent references are shown first.
Citation data come partly from CiteXplore and partly
from an automated harvesting procedure. Note that this is likely to be
only a partial list as not all journals are covered by
either method. However, we are continually building up the citation data
so more and more references will be included with time.
Where a reference describes a PDB structure, the PDB
code is
shown on the right.
|
 |
|




 Links
Links












