 |
PDBsum entry 2ffv

|
|
|
|
|
 |
Contents |
 |
|
|
|
|
|
|
|
|
|
|
|
|
* Residue conservation analysis
|
|
|
|
|
|
|
|
|
|
|
Enzyme class:
|
|
E.C.2.4.1.41
- polypeptide N-acetylgalactosaminyltransferase.
|
|
|
|
|
|
|
Reaction:
|
|
|
1.
|
L-seryl-[protein] + UDP-N-acetyl-alpha-D-galactosamine = a 3-O- [N-acetyl-alpha-D-galactosaminyl]-L-seryl-[protein] + UDP + H+
|
|
2.
|
L-threonyl-[protein] + UDP-N-acetyl-alpha-D-galactosamine = a 3-O- [N-acetyl-alpha-D-galactosaminyl]-L-threonyl-[protein] + UDP + H+
|
|
|
|
|
|
|
L-seryl-[protein]
|
+
|
UDP-N-acetyl-alpha-D-galactosamine
|
=
|
3-O- [N-acetyl-alpha-D-galactosaminyl]-L-seryl-[protein]
|
+
|
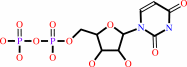 UDP
UDP
|
+
|
H(+)
Bound ligand (Het Group name = )
corresponds exactly
|
|
|
|
|
|
|
L-threonyl-[protein]
|
+
|
UDP-N-acetyl-alpha-D-galactosamine
|
=
|
3-O- [N-acetyl-alpha-D-galactosaminyl]-L-threonyl-[protein]
|
+
|
UDP
|
+
|
H(+)
Bound ligand (Het Group name = )
corresponds exactly
|
|
|
|
|
|
|
|
|
|
Cofactor:
|
|
Ca(2+); Mn(2+)
|
|
|
|
|
|
|
|
|
Molecule diagrams generated from .mol files obtained from the
KEGG ftp site
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
| |
|
|
| |
|
DOI no:
|
J Biol Chem
281:8613-8619
(2006)
|
|
PubMed id:
|
|
|
|
|
|
| |
|
Dynamic association between the catalytic and lectin domains of human UDP-GalNAc:polypeptide alpha-N-acetylgalactosaminyltransferase-2.
|
|
T.A.Fritz,
J.Raman,
L.A.Tabak.
|
|
|
|
|
| |
ABSTRACT
|
|
|
|
| |
|
|
The family of UDP-GalNAc:polypeptide alpha-N-acetylgalactosaminyltransferases
(ppGalNAcTs) is unique among glycosyltransferases, containing both catalytic and
lectin domains that we have previously shown to be closely associated. Here we
describe the x-ray crystal structures of human ppGalNAcT-2 (hT2) bound to the
product UDP at 2.75 A resolution and to UDP and an acceptor peptide substrate
EA2 (PTTDSTTPAPTTK) at 1.64 A resolution. The conformations of both UDP and
residues Arg362-Ser372 vary greatly between the two structures. In the
hT2-UDP-EA2 complex, residues Arg362-Ser373 comprise a loop that forms a lid
over UDP, sealing it in the active site, whereas in the hT2-UDP complex this
loop is folded back, exposing UDP to bulk solvent. EA2 binds in a shallow groove
with threonine 7 positioned consistent with in vitro data showing it to be the
preferred site of glycosylation. The relative orientations of the hT2 catalytic
and lectin domains differ dramatically from that of murine ppGalNAcT-1 and also
vary considerably between the two hT2 complexes. Indeed, in the hT2-UDP-EA2
complex essentially no contact is made between the catalytic and lectin domains
except for the peptide bridge between them. Thus, the hT2 structures reveal an
unexpected flexibility between the catalytic and lectin domains and suggest a
new mechanism used by hT2 to capture glycosylated substrates. Kinetic analysis
of hT2 lacking the lectin domain confirmed the importance of this domain in
acting on glycopeptide but not peptide substrates. The structure of the
hT2-UDP-EA2 complex also resolves long standing questions regarding ppGalNAcT
acceptor substrate specificity.
|
|
|
|
|
|
| |
Selected figure(s)
|
|
|
|
| |
|
|

|
|

|
|
Figure 3.
FIGURE 3. Hydrogen bonds and hydrophobic interactions
mediating binding between hT2 and EA2. EA2 is shown as a
stylized drawing with yellow carbon atoms, and the individual
residues Ser^5–Lys^13 are labeled in red. Hydrogen bonds are
shown by the blue dashed lines along with their corresponding
lengths in Ångstroms. Hydrophobic interactions are shown
by the red "eyelashes." Water molecules are shown as red
spheres. The diagram was created by editing the output from the
program Ligplot (47).
|
|
Figure 4.
FIGURE 4. Stereo view of EA2 binding to hT2. The
transparent surface of hT2 is colored cyan, except for the
surface of flexible loop residues Arg^362–Ser^372, which is
colored yellow. A ribbon diagram of residues Arg^362–Ser^373
is shown in yellow. EA2 is shown with white carbons, and
individual residues are indicated by white letter/number
combinations. The side chains of hT2 residues interacting with
EA2 are indicated by the black letter/number combinations. The
five water molecules in the putative GalNAc binding pocket are
shown as red spheres, only two of which are indicated for
purposes of clarity. Two additional water molecules bound to
shallow pockets in the EA2 cleft are also shown.
|
|
|
|
|
|
| |
The above figures are
reprinted
by permission from the ASBMB:
J Biol Chem
(2006,
281,
8613-8619)
copyright 2006.
|
|
| |
Figures were
selected
by an automated process.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
 |
 |
|
Literature references that cite this PDB file's key reference
|
|
|
| |
PubMed id
|
|
Reference
|
|
|
|
|
|
D.J.Gill,
H.Clausen,
and
F.Bard
(2011).
Location, location, location: new insights into O-GalNAc protein glycosylation.
|
| |
Trends Cell Biol,
21,
149-158.
|
|
|
|
|
|
|
Y.Cai,
J.He,
and
L.Lu
(2011).
Prediction of mucin-type O-glycosylation sites by a two-staged strategy.
|
| |
Mol Divers,
15,
427-433.
|
|
|
|
|
|
|
B.Ramakrishnan,
and
P.K.Qasba
(2010).
Structure-based evolutionary relationship of glycosyltransferases: a case study of vertebrate β1,4-galactosyltransferase, invertebrate β1,4-N-acetylgalactosaminyltransferase and α-polypeptidyl-N-acetylgalactosaminyltransferase.
|
| |
Curr Opin Struct Biol,
20,
536-542.
|
|
|
|
|
|
|
H.E.Miwa,
T.A.Gerken,
O.Jamison,
and
L.A.Tabak
(2010).
Isoform-specific O-glycosylation of osteopontin and bone sialoprotein by polypeptide N-acetylgalactosaminyltransferase-1.
|
| |
J Biol Chem,
285,
1208-1219.
|
|
|
|
|
|
|
R.Hurtado-Guerrero,
T.Zusman,
S.Pathak,
A.F.Ibrahim,
S.Shepherd,
A.Prescott,
G.Segal,
and
D.M.van Aalten
(2010).
Molecular mechanism of elongation factor 1A inhibition by a Legionella pneumophila glycosyltransferase.
|
| |
Biochem J,
426,
281-292.
|
|
|
PDB codes:
|
|
|
|
|
|
|
|
K.Guda,
H.Moinova,
J.He,
O.Jamison,
L.Ravi,
L.Natale,
J.Lutterbaugh,
E.Lawrence,
S.Lewis,
J.K.Willson,
J.B.Lowe,
G.L.Wiesner,
G.Parmigiani,
J.Barnholtz-Sloan,
D.W.Dawson,
V.E.Velculescu,
K.W.Kinzler,
N.Papadopoulos,
B.Vogelstein,
J.Willis,
T.A.Gerken,
and
S.D.Markowitz
(2009).
Inactivating germ-line and somatic mutations in polypeptide N-acetylgalactosaminyltransferase 12 in human colon cancers.
|
| |
Proc Natl Acad Sci U S A,
106,
12921-12925.
|
|
|
|
|
|
|
N.Heise,
D.Singh,
H.van der Wel,
S.O.Sassi,
J.M.Johnson,
C.L.Feasley,
C.M.Koeller,
J.O.Previato,
L.Mendonça-Previato,
and
C.M.West
(2009).
Molecular analysis of a UDP-GlcNAc:polypeptide alpha-N-acetylglucosaminyltransferase implicated in the initiation of mucin-type O-glycosylation in Trypanosoma cruzi.
|
| |
Glycobiology,
19,
918-933.
|
|
|
|
|
|
|
J.Raman,
T.A.Fritz,
T.A.Gerken,
O.Jamison,
D.Live,
M.Liu,
and
L.A.Tabak
(2008).
The catalytic and lectin domains of UDP-GalNAc:polypeptide alpha-N-Acetylgalactosaminyltransferase function in concert to direct glycosylation site selection.
|
| |
J Biol Chem,
283,
22942-22951.
|
|
|
|
|
|
|
L.L.Lairson,
B.Henrissat,
G.J.Davies,
and
S.G.Withers
(2008).
Glycosyltransferases: structures, functions, and mechanisms.
|
| |
Annu Rev Biochem,
77,
521-555.
|
|
|
|
|
|
|
P.K.Qasba,
E.Boeggeman,
and
B.Ramakrishnan
(2008).
Site-specific linking of biomolecules via glycan residues using glycosyltransferases.
|
| |
Biotechnol Prog,
24,
520-526.
|
|
|
|
|
|
|
T.A.Gerken,
K.G.Hagen,
and
O.Jamison
(2008).
Conservation of peptide acceptor preferences between Drosophila and mammalian polypeptide-GalNAc transferase ortholog pairs.
|
| |
Glycobiology,
18,
861-870.
|
|
|
|
|
|
|
A.L.Milac,
N.V.Buchete,
T.A.Fritz,
G.Hummer,
and
L.A.Tabak
(2007).
Substrate-induced conformational changes and dynamics of UDP-N-acetylgalactosamine:polypeptide N-acetylgalactosaminyltransferase-2.
|
| |
J Mol Biol,
373,
439-451.
|
|
|
|
|
|
|
M.R.Bond,
and
J.J.Kohler
(2007).
Chemical methods for glycoprotein discovery.
|
| |
Curr Opin Chem Biol,
11,
52-58.
|
|
|
|
|
|
|
M.Tenno,
A.Saeki,
A.P.Elhammer,
and
A.Kurosaka
(2007).
Function of conserved aromatic residues in the Gal/GalNAc-glycosyltransferase motif of UDP-GalNAc:polypeptide N-acetylgalactosaminyltransferase 1.
|
| |
FEBS J,
274,
6037-6045.
|
|
|
|
|
|
The most recent references are shown first.
Citation data come partly from CiteXplore and partly
from an automated harvesting procedure. Note that this is likely to be
only a partial list as not all journals are covered by
either method. However, we are continually building up the citation data
so more and more references will be included with time.
Where a reference describes a PDB structure, the PDB
codes are
shown on the right.
|
|



 Links
Links














