 |
PDBsum entry 2blc

|
|
|
|
|
|

|

|

|

|
|

|
|

|
|

|
|

|
|
|
 |

|

|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Oxidoreductase
|
PDB id
|
|
|
|
2blc
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|

|

|

|
|
|
|
|
|
|
|
 |
Contents |
 |
|
|
|
|
|
|
|
|
|
|
|
|
* Residue conservation analysis
|
|
|
|
|
|
|
|
|
|
|
Enzyme class 2:
|
|
E.C.1.5.1.3
- dihydrofolate reductase.
|
|
|
|
|
|
|
Pathway:
|
|
Folate Coenzymes
|
|
|
|
|
|
Reaction:
|
|
(6S)-5,6,7,8-tetrahydrofolate + NADP+ = 7,8-dihydrofolate + NADPH + H+
|
|
|
|
|
|
(6S)-5,6,7,8-tetrahydrofolate
|
+
|
NADP(+)
Bound ligand (Het Group name = )
corresponds exactly
|
=
|
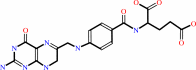 7,8-dihydrofolate
7,8-dihydrofolate
|
+
|
 NADPH
NADPH
|
+
|
H(+)
|
|
|
|
|
|
|
|
|
|
Enzyme class 3:
|
|
E.C.2.1.1.45
- thymidylate synthase.
|
|
|
|
|
|
|
Pathway:
|
|
|
|
|
|
|
|
Reaction:
|
|
dUMP + (6R)-5,10-methylene-5,6,7,8-tetrahydrofolate = 7,8-dihydrofolate + dTMP
|
|
|
|
|
|
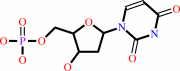 dUMP
dUMP
|
+
|
(6R)-5,10-methylene-5,6,7,8-tetrahydrofolate
Bound ligand (Het Group name = )
matches with 44.00% similarity
|
=
|
7,8-dihydrofolate
|
+
|
dTMP
Bound ligand (Het Group name = )
matches with 40.82% similarity
|
|
|
|
|
|
|
|
|
|
|
|
|
Note, where more than one E.C. class is given (as above), each may
correspond to a different protein domain or, in the case of polyprotein
precursors, to a different mature protein.
|
|
|
|
Molecule diagrams generated from .mol files obtained from the
KEGG ftp site
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
| |
|
|
| |
|
DOI no:
|
Proc Natl Acad Sci U S A
102:13046-13051
(2005)
|
|
PubMed id:
|
|
|
|
|
|
| |
|
Crystal structure of dihydrofolate reductase from Plasmodium vivax: pyrimethamine displacement linked with mutation-induced resistance.
|
|
P.Kongsaeree,
P.Khongsuk,
U.Leartsakulpanich,
P.Chitnumsub,
B.Tarnchompoo,
M.D.Walkinshaw,
Y.Yuthavong.
|
|
|
|
|
| |
ABSTRACT
|
|
|
|
| |
|
|
Pyrimethamine (Pyr) targets dihydrofolate reductase of Plasmodium vivax (PvDHFR)
as well as other malarial parasites, but its use as antimalarial is hampered by
the widespread high resistance. Comparison of the crystal structures of PvDHFR
from wild-type and the Pyr-resistant (SP21, Ser-58 --> Arg + Ser-117 --> Asn)
strain as complexes with NADPH and Pyr or its analog lacking p-Cl (Pyr20)
clearly shows that the steric conflict arising from the side chain of Asn-117 in
the mutant enzyme, accompanied by the loss of binding to Ser-120, is mainly
responsible for the reduction in binding of Pyr. Pyr20 still effectively
inhibits both the wild-type and SP21 proteins, and the x-ray structures of these
complexes show how Pyr20 fits into both active sites without steric strain.
These structural insights suggest a general approach for developing new
generations of antimalarial DHFR inhibitors that, by only occupying substrate
space of the active site, would retain binding affinity with the mutant enzymes.
|
|
|
|
|
|
| |
Selected figure(s)
|
|
|
|
| |
|
|

|
|

|
|
Figure 1.
Fig. 1. Crystal structure of the DHFR domain of P. vivax.
(A) Structure of P. vivax DHFR complexed with NADPH and Pyr;
 -helices are in red,
and -helices are in red,
and  -strands are in blue,
including the Insert-1 loop and the Insert-2 -helix. The carbons,
nitrogen, oxygen, and chlorine atoms of Pyr and NADPH are shown
in yellow, blue, red, and magenta, respectively. (B) Comparison
of the DHFR domains from P. vivax (green) and P. falciparum
(magenta). The superimposed structures demonstrate overall
structural similarity with major deviation in the insert regions. -strands are in blue,
including the Insert-1 loop and the Insert-2 -helix. The carbons,
nitrogen, oxygen, and chlorine atoms of Pyr and NADPH are shown
in yellow, blue, red, and magenta, respectively. (B) Comparison
of the DHFR domains from P. vivax (green) and P. falciparum
(magenta). The superimposed structures demonstrate overall
structural similarity with major deviation in the insert regions.
|
|
Figure 2.
Fig. 2. Pyr bound in the PvDHFR active site. The Pyr and
NADPH cofactor are shown as balls and sticks with carbon,
nitrogen, and chlorine colored yellow, blue, and magenta,
respectively. (A) Pyr binding with the WT PvDHFR. Interactions
between the enzyme and the pyrimidine ring of the inhibitor
include electrostatic interactions and H-bonds indicated by
dotted lines. Numbers next to the lines indicate distances in
Å. (B) Pyr binding with the SP21 double-mutant PvDHFR.
Interactions around the pyrimidine ring are similar to the WT
enzyme. The mutation at codon 117 from Ser to Asn increases a
steric factor in the active site, and, as a result, the
positions of both NADPH and Pyr are perturbed from their optimum
binding, reducing the efficiency of Pyr by as much as 300-fold.
(C) Superposition of Pyr-binding sites in the WT PvDHFR (green)
and the SP21 double-mutant enzyme (orange) with a rmsd of 0.53
Å. While the position of pyrimidine is held in the same
place, the mutation at S117N causes the displacement of
p-chlorophenyl moiety of Pyr from its optimal binding with the
p-Cl atom shifted by 1.1 Å and the torsion plane between
the two rings twisted by -32°. The mutation also caused a
local main-chain movement of residues 118 -125 (0.60 -1.88
Å), with respect to the WT enzyme. The mutation at codon
58 from Ser to Arg is not in proximity with the Pyr-binding site.
|
|
|
|
| |
Figures were
selected
by an automated process.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
 |
 |
|
Literature references that cite this PDB file's key reference
|
|
|
| |
PubMed id
|
|
Reference
|
|
|
|
|
|
A.M.Auliff,
J.H.Adams,
M.T.O'Neil,
and
Q.Cheng
(2010).
Defining the role of mutations in Plasmodium vivax dihydrofolate reductase-thymidylate synthase gene using an episomal Plasmodium falciparum transfection system.
|
| |
Antimicrob Agents Chemother,
54,
3927-3932.
|
|
|
|
|
|
|
K.Choowongkomon,
S.Theppabutr,
N.Songtawee,
N.P.Day,
N.J.White,
C.J.Woodrow,
and
M.Imwong
(2010).
Computational analysis of binding between malarial dihydrofolate reductases and anti-folates.
|
| |
Malar J,
9,
65.
|
|
|
|
|
|
|
C.R.Bourne,
R.A.Bunce,
P.C.Bourne,
K.D.Berlin,
E.W.Barrow,
and
W.W.Barrow
(2009).
Crystal structure of Bacillus anthracis dihydrofolate reductase with the dihydrophthalazine-based trimethoprim derivative RAB1 provides a structural explanation of potency and selectivity.
|
| |
Antimicrob Agents Chemother,
53,
3065-3073.
|
|
|
PDB codes:
|
|
|
|
|
|
|
|
P.Sivaprakasam,
P.N.Tosso,
and
R.J.Doerksen
(2009).
Structure-activity relationship and comparative docking studies for cycloguanil analogs as PfDHFR-TS inhibitors.
|
| |
J Chem Inf Model,
49,
1787-1796.
|
|
|
|
|
|
|
V.Kairys,
M.K.Gilson,
V.Lather,
C.A.Schiffer,
and
M.X.Fernandes
(2009).
Toward the design of mutation-resistant enzyme inhibitors: further evaluation of the substrate envelope hypothesis.
|
| |
Chem Biol Drug Des,
74,
234-245.
|
|
|
|
|
|
|
V.Kasam,
J.Salzemann,
M.Botha,
A.Dacosta,
G.Degliesposti,
R.Isea,
D.Kim,
A.Maass,
C.Kenyon,
G.Rastelli,
M.Hofmann-Apitius,
and
V.Breton
(2009).
WISDOM-II: Screening against multiple targets implicated in malaria using computational grid infrastructures.
|
| |
Malar J,
8,
88.
|
|
|
|
|
|
|
D.Hecht,
M.Cheung,
and
G.B.Fogel
(2008).
QSAR using evolved neural networks for the inhibition of mutant PfDHFR by pyrimethamine derivatives.
|
| |
Biosystems,
92,
10-15.
|
|
|
|
|
|
|
G.B.Fogel,
M.Cheung,
E.Pittman,
and
D.Hecht
(2008).
Modeling the inhibition of quadruple mutant Plasmodium falciparum dihydrofolate reductase by pyrimethamine derivatives.
|
| |
J Comput Aided Mol Des,
22,
29-38.
|
|
|
|
|
|
|
J.E.Hyde
(2007).
Targeting purine and pyrimidine metabolism in human apicomplexan parasites.
|
| |
Curr Drug Targets,
8,
31-47.
|
|
|
|
|
|
|
A.Argyrou,
M.W.Vetting,
B.Aladegbami,
and
J.S.Blanchard
(2006).
Mycobacterium tuberculosis dihydrofolate reductase is a target for isoniazid.
|
| |
Nat Struct Mol Biol,
13,
408-413.
|
|
|
PDB code:
|
|
|
|
|
|
|
|
L.Y.Djapa,
R.Zelikson,
A.Delahodde,
M.Bolotin-Fukuhara,
and
A.Mazabraud
(2006).
Plasmodium vivax dihydrofolate reductase as a target of sulpha drugs.
|
| |
FEMS Microbiol Lett,
256,
105-111.
|
|
|
|
|
|
|
S.Bunyarataphan,
U.Leartsakulpanich,
S.Taweechai,
B.Tarnchompoo,
S.Kamchonwongpaisan,
and
Y.Yuthavong
(2006).
Evaluation of the activities of pyrimethamine analogs against Plasmodium vivax and Plasmodium falciparum dihydrofolate reductase-thymidylate synthase using in vitro enzyme inhibition and bacterial complementation assays.
|
| |
Antimicrob Agents Chemother,
50,
3631-3637.
|
|
|
|
|
|
|
V.Cody,
J.Pace,
K.Chisum,
and
A.Rosowsky
(2006).
New insights into DHFR interactions: analysis of Pneumocystis carinii and mouse DHFR complexes with NADPH and two highly potent 5-(omega-carboxy(alkyloxy) trimethoprim derivatives reveals conformational correlations with activity and novel parallel ring stacking interactions.
|
| |
Proteins,
65,
959-969.
|
|
|
PDB codes:
|
|
|
|
|
|
|
|
Y.Yuthavong,
S.Kamchonwongpaisan,
U.Leartsakulpanich,
and
P.Chitnumsub
(2006).
Folate metabolism as a source of molecular targets for antimalarials.
|
| |
Future Microbiol,
1,
113-125.
|
|
|
|
|
|
The most recent references are shown first.
Citation data come partly from CiteXplore and partly
from an automated harvesting procedure. Note that this is likely to be
only a partial list as not all journals are covered by
either method. However, we are continually building up the citation data
so more and more references will be included with time.
Where a reference describes a PDB structure, the PDB
codes are
shown on the right.
|
|


 Links
Links













