 |
PDBsum entry 1eq3

|
|
|
|
|
 |
Contents |
 |
|
|
|
|
|
|
|
|
* Residue conservation analysis
|
|
|
|
|
|
|
|
|
|
|
Enzyme class:
|
|
E.C.5.2.1.8
- peptidylprolyl isomerase.
|
|
|
|
|
|
|
Reaction:
|
|
[protein]-peptidylproline (omega=180) = [protein]-peptidylproline (omega=0)
|
|
|
|
|
|
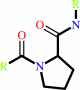 Peptidylproline (omega=180)
Peptidylproline (omega=180)
|
=
|
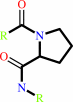 peptidylproline (omega=0)
peptidylproline (omega=0)
|
|
|
|
|
|
|
|
|
|
|
|
|
Molecule diagrams generated from .mol files obtained from the
KEGG ftp site
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
| |
|
|
| |
|
DOI no:
|
J Mol Biol
301:1003-1017
(2000)
|
|
PubMed id:
|
|
|
|
|
|
| |
|
NMR solution structure of hPar14 reveals similarity to the peptidyl prolyl cis/trans isomerase domain of the mitotic regulator hPin1 but indicates a different functionality of the protein.
|
|
E.Sekerina,
J.U.Rahfeld,
J.Müller,
J.Fanghänel,
C.Rascher,
G.Fischer,
P.Bayer.
|
|
|
|
|
| |
ABSTRACT
|
|
|
|
| |
|
|
The 131-amino acid residue parvulin-like human peptidyl-prolyl cis/trans
isomerase (PPIase) hPar14 was shown to exhibit sequence similarity to the
regulator enzyme for cell cycle transitions human hPin1, but specificity for
catalyzing pSer(Thr)-Pro cis/trans isomerizations was lacking. To determine the
solution structure of hPar14 the (1)H, (13)C, and (15)N chemical shifts of this
protein have been assigned using heteronuclear two and three-dimensional NMR
experiments on unlabeled and uniformly (15)N/(13)C-labeled recombinant protein
isolated from Escherichia coli cells that overexpress the protein. The chemical
shift assignments were used to interpret the NOE data, which resulted in a total
of 1042 NOE restraints. The NOE restraints were used along with 71 dihedral
angle restraints and 38 hydrogen bonding restraints to produce 50 low-energy
structures. The hPar14 folds into a betaalpha(3)betaalphabeta(2) structure, and
contains an unstructured 35-amino acid basic tail N-terminal to the catalytic
core that replaces the WW domain of hPin1 homologs. The three-dimensional
structures of hPar14 and the PPIase domain of human hPin1 reveal a high degree
of conservation. The root-mean-square deviations of the mean atomic coordinates
of the heavy atoms of the backbone between residues 38 to 45, 50 to 58, 64 to
70, 81 to 86, 115 to 119 and 122 to 128 of hPar14 were 0.81(+/-0.07) A. The
hPar14 model structure provides insight into how this class of PPIases may
select preferential secondary catalytic sites, and also allows identification of
a putative DNA-binding motif in parvulin-like PPIases.
|
|
|
|
|
|
| |
Selected figure(s)
|
|
|
|
| |
|
|

|
|

|
|
Figure 7.
Figure 7. Overlay of the active center of hPin1 (blue) and
hPar14 (red). Superposition of the putative active site and the
supposed catalytic active amino acid residues in both human
parvulins. The gray ribbon represents the backbone fold of
hPar14.
|
|
Figure 9.
Figure 9. GRASP representation of molecular surfaces of
hPin1 and hPar14^36-131. The electrostatic surface potential is
indicated by colours (red, acidic; blue, basic). (a) View on the
surface along the active center of hPar14. Conserved residues
are labeled in red, yellow, green or magenta. Color codes mark
residues from which counterparts can be found at simular spatial
positions in hPin1 (d). (b) Surface backside of hPar14. The
quadruple of residues forming two salt bridges are labeled. (c)
Negative spot surrounded by hydrophobic side-chains on the
surface of hPar14. The acidic region is formed by amino acid
residues of the loop region from Pro102 to Pro109, the following
b-strand Val110 to Thr112 and the conserved turn Asp113 to
Pro115. (d) View on the surface along the active center of
hPin1. Color codes used as in (a).
|
|
|
|
|
|
| |
The above figures are
reprinted
by permission from Elsevier:
J Mol Biol
(2000,
301,
1003-1017)
copyright 2000.
|
|
| |
Figures were
selected
by an automated process.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
 |
 |
|
Literature references that cite this PDB file's key reference
|
|
|
| |
PubMed id
|
|
Reference
|
|
|
|
|
|
L.Martino,
Y.He,
K.L.Hands-Taylor,
E.R.Valentine,
G.Kelly,
C.Giancola,
and
M.R.Conte
(2009).
The interaction of the Escherichia coli protein SlyD with nickel ions illuminates the mechanism of regulation of its peptidyl-prolyl isomerase activity.
|
| |
FEBS J,
276,
4529-4544.
|
|
|
PDB code:
|
|
|
|
|
|
|
|
O.Heikkinen,
R.Seppala,
H.Tossavainen,
S.Heikkinen,
H.Koskela,
P.Permi,
and
I.Kilpeläinen
(2009).
Solution structure of the parvulin-type PPIase domain of Staphylococcus aureus PrsA--implications for the catalytic mechanism of parvulins.
|
| |
BMC Struct Biol,
9,
17.
|
|
|
PDB code:
|
|
|
|
|
|
|
|
S.Fujiyama-Nakamura,
H.Yoshikawa,
K.Homma,
T.Hayano,
T.Tsujimura-Takahashi,
K.Izumikawa,
H.Ishikawa,
N.Miyazawa,
M.Yanagida,
Y.Miura,
T.Shinkawa,
Y.Yamauchi,
T.Isobe,
and
N.Takahashi
(2009).
Parvulin (Par14), a peptidyl-prolyl cis-trans isomerase, is a novel rRNA processing factor that evolved in the metazoan lineage.
|
| |
Mol Cell Proteomics,
8,
1552-1565.
|
|
|
|
|
|
|
J.W.Mueller,
and
P.Bayer
(2008).
Small family with key contacts: par14 and par17 parvulin proteins, relatives of pin1, now emerge in biomedical research.
|
| |
Perspect Medicin Chem,
2,
11-20.
|
|
|
|
|
|
|
V.Lamour,
S.T.Rutherford,
K.Kuznedelov,
U.A.Ramagopal,
R.L.Gourse,
K.Severinov,
and
S.A.Darst
(2008).
Crystal structure of Escherichia coli Rnk, a new RNA polymerase-interacting protein.
|
| |
J Mol Biol,
383,
367-379.
|
|
|
PDB code:
|
|
|
|
|
|
|
|
D.Kessler,
P.Papatheodorou,
T.Stratmann,
E.A.Dian,
C.Hartmann-Fatu,
J.Rassow,
P.Bayer,
and
J.W.Mueller
(2007).
The DNA binding parvulin Par17 is targeted to the mitochondrial matrix by a recently evolved prepeptide uniquely present in Hominidae.
|
| |
BMC Biol,
5,
37.
|
|
|
|
|
|
|
J.W.Mueller,
D.Kessler,
D.Neumann,
T.Stratmann,
P.Papatheodorou,
C.Hartmann-Fatu,
and
P.Bayer
(2006).
Characterization of novel elongated Parvulin isoforms that are ubiquitously expressed in human tissues and originate from alternative transcription initiation.
|
| |
BMC Mol Biol,
7,
9.
|
|
|
|
|
|
|
A.Kühlewein,
G.Voll,
B.Hernandez Alvarez,
H.Kessler,
G.Fischer,
J.U.Rahfeld,
and
G.Gemmecker
(2004).
Solution structure of Escherichia coli Par10: The prototypic member of the Parvulin family of peptidyl-prolyl cis/trans isomerases.
|
| |
Protein Sci,
13,
2378-2387.
|
|
|
PDB codes:
|
|
|
|
|
|
|
|
M.Vitikainen,
I.Lappalainen,
R.Seppala,
H.Antelmann,
H.Boer,
S.Taira,
H.Savilahti,
M.Hecker,
M.Vihinen,
M.Sarvas,
and
V.P.Kontinen
(2004).
Structure-function analysis of PrsA reveals roles for the parvulin-like and flanking N- and C-terminal domains in protein folding and secretion in Bacillus subtilis.
|
| |
J Biol Chem,
279,
19302-19314.
|
|
|
|
|
|
|
D.M.Jacobs,
K.Saxena,
M.Vogtherr,
P.Bernado,
M.Pons,
and
K.M.Fiebig
(2003).
Peptide binding induces large scale changes in inter-domain mobility in human Pin1.
|
| |
J Biol Chem,
278,
26174-26182.
|
|
|
|
|
|
|
T.Uchida,
M.Takamiya,
M.Takahashi,
H.Miyashita,
H.Ikeda,
T.Terada,
Y.Matsuo,
M.Shirouzu,
S.Yokoyama,
F.Fujimori,
and
T.Hunter
(2003).
Pin1 and Par14 peptidyl prolyl isomerase inhibitors block cell proliferation.
|
| |
Chem Biol,
10,
15-24.
|
|
|
|
|
|
|
E.Bitto,
and
D.B.McKay
(2002).
Crystallographic structure of SurA, a molecular chaperone that facilitates folding of outer membrane porins.
|
| |
Structure,
10,
1489-1498.
|
|
|
PDB code:
|
|
|
|
|
|
|
|
S.Fujiyama,
M.Yanagida,
T.Hayano,
Y.Miura,
T.Isobe,
F.Fujimori,
T.Uchida,
and
N.Takahashi
(2002).
Isolation and proteomic characterization of human Parvulin-associating preribosomal ribonucleoprotein complexes.
|
| |
J Biol Chem,
277,
23773-23780.
|
|
|
|
|
|
The most recent references are shown first.
Citation data come partly from CiteXplore and partly
from an automated harvesting procedure. Note that this is likely to be
only a partial list as not all journals are covered by
either method. However, we are continually building up the citation data
so more and more references will be included with time.
Where a reference describes a PDB structure, the PDB
code is
shown on the right.
|
|


 Links
Links











