 |
PDBsum entry 1euq

|
|
|
|
|
 |
Contents |
 |
|
|
|
|
|
|
|
|
|
|
|
|
* Residue conservation analysis
|
|
|
|
|
|
PDB id:
|
|
|
|
| Name: |
|
Ligase/RNA
|
|
|
Title:
|
|
Crystal structure of glutaminyl-tRNA synthetase complexed with a tRNA- gln mutant and an active-site inhibitor
|
|
Structure:
|
|
Glutaminyl tRNA. Chain: b. Engineered: yes. Glutaminyl-tRNA synthetase. Chain: a. Synonym: glnrs, glutamine-tRNA ligase. Engineered: yes
|
|
Source:
|
|
Synthetic: yes. Other_details: product of runoff t7 polymerase transcription from synthetic DNA template. Escherichia coli. Organism_taxid: 562. Expressed in: escherichia coli. Expression_system_taxid: 562
|
|
Biol. unit:
|
|
Dimer (from
)
|
|
Resolution:
|
|
|
3.10Å
|
R-factor:
|
0.242
|
R-free:
|
0.285
|
|
|
Authors:
|
|
L.D.Sherlin,T.L.Bullock,K.J.Newberry,R.S.A.Lipman,Y.-M.Hou,B.Beijer, B.S.Sproat,J.J.Perona
|
Key ref:

|
|
L.D.Sherlin
et al.
(2000).
Influence of transfer RNA tertiary structure on aminoacylation efficiency by glutaminyl and cysteinyl-tRNA synthetases.
J Mol Biol,
299,
431-446.
PubMed id:
DOI:

|
|
|
Date:
|
|
|
17-Apr-00
|
Release date:
|
04-Jun-00
|
|
|
|
|
|
|
PROCHECK
|

|
|
|
|
|
Headers
|
|
|
|
References
|
|
|
|
|
|

|
|
P00962
(SYQ_ECOLI) -
Glutamine--tRNA ligase from Escherichia coli (strain K12)
|
|
|
|
Seq:
Struc:
|
|
|
|
554 a.a.
529 a.a.
|
|

|

|
|
|
|
|
|
|
|
|
|
|
|
Key: |
 |
PfamA domain |
|
|
 |
Secondary structure |
|
 |
CATH domain |
|
|
|
|
|
|
|
|

|
|
|
|
G-G-G-G-U-A-U-C-G-C-C-A-A-G-C-G-G-U-A-A-G-G-C-A-C-C-G-G-A-U-U-C-U-G-A-U-U-C-C-
72 bases
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Enzyme class:
|
|
E.C.6.1.1.18
- glutamine--tRNA ligase.
|
|
|
|
|
|
|
Reaction:
|
|
tRNA(Gln) + L-glutamine + ATP = L-glutaminyl-tRNA(Gln) + AMP + diphosphate
|
|
|
|
|
|
 tRNA(Gln)
tRNA(Gln)
|
+
|
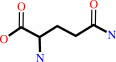 L-glutamine
L-glutamine
|
+
|
 ATP
ATP
|
=
|
L-glutaminyl-tRNA(Gln)
Bound ligand (Het Group name = )
matches with 52.78% similarity
|
+
|
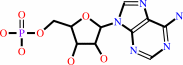 AMP
AMP
|
+
|
 diphosphate
diphosphate
|
|
|
|
|
|
|
|
|
|
|
|
|
Molecule diagrams generated from .mol files obtained from the
KEGG ftp site
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
| |
|
|
| |
|
DOI no:
|
J Mol Biol
299:431-446
(2000)
|
|
PubMed id:
|
|
|
|
|
|
| |
|
Influence of transfer RNA tertiary structure on aminoacylation efficiency by glutaminyl and cysteinyl-tRNA synthetases.
|
|
L.D.Sherlin,
T.L.Bullock,
K.J.Newberry,
R.S.Lipman,
Y.M.Hou,
B.Beijer,
B.S.Sproat,
J.J.Perona.
|
|
|
|
|
| |
ABSTRACT
|
|
|
|
| |
|
|
The position of the tertiary Levitt pair between nucleotides 15 and 48 in the
transfer RNA core region suggests a key role in stabilizing the joining of the
two helical domains, and in maintaining the relative orientations of the D and
variable loops. E. coli tRNA(Gln) possesses the canonical Pu15-Py48 trans
pairing at this position (G15-C48), while the tRNA(Cys) species from this
organism instead features an unusual G15-G48 pair. To explore the structural
context dependence of a G15-G48 Levitt pair, a number of tRNA(Gln) species
containing G15-G48 were constructed and evaluated as substrates for glutaminyl
and cysteinyl-tRNA synthetases. The glutaminylation efficiencies of these mutant
tRNAs are reduced by two to tenfold compared with native tRNA(Gln), consistent
with previous findings that the tertiary core of this tRNA plays a role in GlnRS
recognition. Introduction of tRNA(Cys) identity nucleotides at the acceptor and
anticodon ends of tRNA(Gln) produced a tRNA substrate which was efficiently
aminoacylated by CysRS, even though the tertiary core region of this species
contains the tRNA(Gln) G15-C48 pair. Surprisingly, introduction of G15-G48 into
the non-cognate tRNA(Gln) tertiary core then significantly impairs CysRS
recognition. By contrast, previous work has shown that CysRS aminoacylates
tRNA(Cys) core regions containing G15-G48 with much better efficiency than those
with G15-C48. Therefore, tertiary nucleotides surrounding the Levitt pair must
significantly modulate the efficiency of aminoacylation by CysRS. To explore the
detailed nature of the structural interdependence, crystal structures of two
tRNA(Gln) mutants containing G15-G48 were determined bound to GlnRS. These
structures show that the larger purine ring of G48 is accommodated by rotation
into the syn position, with the N7 nitrogen serving as hydrogen bond acceptor
from several groups of G15. The G15-G48 conformations differ significantly
compared to that observed in the native tRNA(Cys) structure bound to EF-Tu,
further implicating an important role for surrounding nucleotides in maintaining
the integrity of the tertiary core and its consequent ability to present crucial
recognition determinants to aminoacyl-tRNA synthetases.
|
|
|
|
|
|
| |
Selected figure(s)
|
|
|
|
| |
|
|

|
|

|
|
Figure 6.
Figure 6. Hydrogen bonding interactions of the 15-48 Levitt
pair in tRNA^Gln (top; [Rould et al 1991]), tRNA^Cys (center;
[Nissen et al 1999]) and the Gln04 mutant (bottom). The
distances (in Å) between the electronegative atoms of the
hydrogen-bonding pair are indicated.
|
|
Figure 8.
Figure 8. (a) Superposition of the structures of wild-type
tRNA^Gln (bound to GlnRS and QSI) with the Gln04 mutant, in the
region of the 15-48 Levitt pair. The superposition is based on
backbone atoms of the protein (residues 8-547). The wild-type
structure is shown in blue and green and the Gln04 mutant in red
and yellow. (b) Divergent stereo view of the superposition in
(a) with the same color-coding. Nucleotide U8 of both structures
and nucleotides U46 and U47 of wild-type are removed for
clarity. The new tertiary hydrogen bond formed in the Gln04
mutant is indicated by the broken black line.
|
|
|
|
|
|
| |
The above figures are
reprinted
by permission from Elsevier:
J Mol Biol
(2000,
299,
431-446)
copyright 2000.
|
|
| |
Figures were
selected
by an automated process.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
 |
 |
|
Literature references that cite this PDB file's key reference
|
|
|
| |
PubMed id
|
|
Reference
|
|
|
|
|
|
I.A.Vasil'eva,
and
N.A.Moor
(2007).
Interaction of aminoacyl-tRNA synthetases with tRNA: general principles and distinguishing characteristics of the high-molecular-weight substrate recognition.
|
| |
Biochemistry (Mosc),
72,
247-263.
|
|
|
|
|
|
|
L.E.Sanderson,
and
O.C.Uhlenbeck
(2007).
The 51-63 base pair of tRNA confers specificity for binding by EF-Tu.
|
| |
RNA,
13,
835-840.
|
|
|
|
|
|
|
I.Gruic-Sovulj,
N.Uter,
T.Bullock,
and
J.J.Perona
(2005).
tRNA-dependent aminoacyl-adenylate hydrolysis by a nonediting class I aminoacyl-tRNA synthetase.
|
| |
J Biol Chem,
280,
23978-23986.
|
|
|
PDB code:
|
|
|
|
|
|
|
|
S.Hauenstein,
C.M.Zhang,
Y.M.Hou,
and
J.J.Perona
(2004).
Shape-selective RNA recognition by cysteinyl-tRNA synthetase.
|
| |
Nat Struct Mol Biol,
11,
1134-1141.
|
|
|
PDB code:
|
|
|
|
|
|
|
|
C.Evilia,
X.Ming,
S.Dassarma,
and
Y.M.Hou
(2003).
Aminoacylation of an unusual tRNA(Cys) from an extreme halophile.
|
| |
RNA,
9,
794-801.
|
|
|
|
|
|
|
L.D.Sherlin,
T.L.Bullock,
T.A.Nissan,
J.J.Perona,
F.J.Lariviere,
O.C.Uhlenbeck,
and
S.A.Scaringe
(2001).
Chemical and enzymatic synthesis of tRNAs for high-throughput crystallization.
|
| |
RNA,
7,
1671-1678.
|
|
|
|
|
|
|
M.Szymanski,
M.A.Deniziak,
and
J.Barciszewski
(2001).
Aminoacyl-tRNA synthetases database.
|
| |
Nucleic Acids Res,
29,
288-290.
|
|
|
|
|
|
|
T.A.Nissan,
and
J.J.Perona
(2000).
Alternative designs for construction of the class II transfer RNA tertiary core.
|
| |
RNA,
6,
1585-1596.
|
|
|
|
|
|
The most recent references are shown first.
Citation data come partly from CiteXplore and partly
from an automated harvesting procedure. Note that this is likely to be
only a partial list as not all journals are covered by
either method. However, we are continually building up the citation data
so more and more references will be included with time.
Where a reference describes a PDB structure, the PDB
code is
shown on the right.
|
|


 Links
Links






