|
Figure 5.
Figure 5 Hypothetical model of ATP hydrolysis-facilitated gap
displacement. The crystallized protein filament is shown as a C^
 trace
in salmon with AMPPNP in green. Speculative models of the DNA
substrates are shown as wires. Important segments are boxed. The
5'- and 3'-ends are based on the filament-initiating ssDNA
(thinner wire in blue). The homologous dsDNA is shown as a
thicker wire in yellow. The strand-exchange process progresses
from the 5'-end to the 3'-end. (a) An intervening gap. Such gaps
are likely to exist owing to simultaneous homologous pairing
between the recombinase/ssDNA filament and dsDNA at multiple
locations. The dsDNA in the gap region cannot become properly
wound ( trace
in salmon with AMPPNP in green. Speculative models of the DNA
substrates are shown as wires. Important segments are boxed. The
5'- and 3'-ends are based on the filament-initiating ssDNA
(thinner wire in blue). The homologous dsDNA is shown as a
thicker wire in yellow. The strand-exchange process progresses
from the 5'-end to the 3'-end. (a) An intervening gap. Such gaps
are likely to exist owing to simultaneous homologous pairing
between the recombinase/ssDNA filament and dsDNA at multiple
locations. The dsDNA in the gap region cannot become properly
wound ( 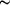 19
bp per helical turn) around the nucleoprotein filament without
unwinding its adjacent region(s). Despite the sequence homology,
it serves as a topological roadblock of strand exchange between
long DNA substrates. (b) ATP hydrolysis promotes the transient
release of a dsDNA segment at the immediate 3'-flank of the gap.
The transiently released dsDNA region is shown as an exaggerated
wide helix. (c) Rearrangement in the transiently released dsDNA
region and the adjacent gap takes place without changing the
overall topology. The 5'-end of the gap region becomes properly
wound, while the released dsDNA region becomes unwound. (d) The
rearranged 5'-end of the gap becomes bound by the recombinase
filament. As a result, the gap is displaced towards the 3'-end.
(e) Repetition of steps (b)-(d) would chase the topologically
strained gap out of the 3'-end of the nucleoprotein filament,
therefore removing topological roadblocks to extensive DNA
strand exchange. 19
bp per helical turn) around the nucleoprotein filament without
unwinding its adjacent region(s). Despite the sequence homology,
it serves as a topological roadblock of strand exchange between
long DNA substrates. (b) ATP hydrolysis promotes the transient
release of a dsDNA segment at the immediate 3'-flank of the gap.
The transiently released dsDNA region is shown as an exaggerated
wide helix. (c) Rearrangement in the transiently released dsDNA
region and the adjacent gap takes place without changing the
overall topology. The 5'-end of the gap region becomes properly
wound, while the released dsDNA region becomes unwound. (d) The
rearranged 5'-end of the gap becomes bound by the recombinase
filament. As a result, the gap is displaced towards the 3'-end.
(e) Repetition of steps (b)-(d) would chase the topologically
strained gap out of the 3'-end of the nucleoprotein filament,
therefore removing topological roadblocks to extensive DNA
strand exchange.
|
