 |
PDBsum entry 3dqh

|
|
|
|
|
|

|
|

|
|

|
|

|
|

|
|
|
 |

|

|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Luminescent protein
|
PDB id
|
|
|
|
3dqh
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|

|

|

|
|
|
|
|
|
References listed in PDB file
|
|
|
Key reference
|
|
|
Title
|
|
Alteration of citrine structure by hydrostatic pressure explains the accompanying spectral shift.
|
|
|
Authors
|
|
B.Barstow,
N.Ando,
C.U.Kim,
S.M.Gruner.
|
|
|
Ref.
|
|
Proc Natl Acad Sci U S A, 2008,
105,
13362-13366.
[DOI no: ]
|
|
|
PubMed id
|
|
|
|
|
|
Abstract
|
|
|
A protein molecule is an intricate system whose function is highly sensitive to
small external perturbations. However, no examples that correlate protein
function with progressive subangstrom structural perturbations have thus far
been presented. To elucidate this relationship, we have investigated a
fluorescent protein, citrine, as a model system under high-pressure
perturbation. The protein has been compressed to produce deformations of its
chromophore by applying a high-pressure cryocooling technique. A closely spaced
series of x-ray crystallographic structures reveals that the chromophore
undergoes a progressive deformation of up to 0.8 A at an applied pressure of 500
MPa. It is experimentally demonstrated that the structural motion is directly
correlated with the progressive fluorescence shift of citrine from yellow to
green under these conditions. This protein is therefore highly sensitive to
subangstrom deformations and its function must be understood at the subangstrom
level. These results have significant implications for protein function
prediction and biomolecule design and engineering, because they suggest methods
to tune protein function by modification of the protein scaffold.
|
|
|
|

|
|

|
|
Figure 1.
The citrine molecule and citrine's chromophore.
|
|
Figure 4.
Average center of mass positions of the main chromophore
phenol and imidazolinone rings in the coordinate system defined
in Fig. 3. Error bars were estimated by Cruickshank's formula.
Fits are shown as solid gray lines.
|
|
|
|
|
|
Secondary reference #1
|
|
|
Title
|
|
High-Pressure cooling of protein crystals without cryoprotectants.
|
|
|
Authors
|
|
C.U.Kim,
R.Kapfer,
S.M.Gruner.
|
|
|
Ref.
|
|
Acta Crystallogr D Biol Crystallogr, 2005,
61,
881-890.
[DOI no: ]
|
|
|
PubMed id
|
|
|
|
|
|
|
|
|
|

|
|

|
|
Figure 2.
Figure 2 High-pressure cooling apparatus. (a) Sample pin. An
oil-coated crystal in a cryoloop is inserted into one end
(right) of a brass tube (2.5 cm long) and a steel piano wire in
the other end (left). (b) High-pressure tubing assembly in a
LN[2] container. A sample is loaded into the top of each 30 cm
long tube and is held in place by a magnet outside the tube.
Three crystals can be pressure-cooled at a time. (c)
High-pressure tubing assembly in a carbon-steel container. The
assembly is connected to a He-gas compressor (rear). All
high-pressure operations are controlled remotely for safety
reasons.
|
|
Figure 5.
Figure 5 AHP-LAAO. (a) Diffraction image of a crystal
flash-cooled at ambient pressure ( 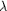 =
0.9795 Å, beam diameter = 150 µm, =
0.9795 Å, beam diameter = 150 µm, 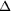 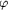 =
1.0°, d = 200 mm, 60 s). Strong ice rings are seen. The
diffraction resolution is only 7.0 Å and the mosaicity is
very poor. (b) Diffraction image of a crystal pressure-cooled at
190 MPa ( =
0.9795 Å, beam diameter = 150 µm, =
1.0°, d = 250 mm, 60 s). The diffraction resolution reaches
2.7 Å and the mosaicity is 0.56°. =
1.0°, d = 200 mm, 60 s). Strong ice rings are seen. The
diffraction resolution is only 7.0 Å and the mosaicity is
very poor. (b) Diffraction image of a crystal pressure-cooled at
190 MPa ( =
0.9795 Å, beam diameter = 150 µm, =
1.0°, d = 250 mm, 60 s). The diffraction resolution reaches
2.7 Å and the mosaicity is 0.56°.
|
|
|
|
|
|
The above figures are
reproduced from the cited reference
with permission from the IUCr
|
|
|
|
|
|
|




