 |
PDBsum entry 3ei7

|
|
|
|
|
 |
Contents |
 |
|
|
|
|
|
|
|
|
|
|
|
|
* Residue conservation analysis
|
|
|
|
|
|
PDB id:
|
|
|
|
| Name: |
|
Transferase
|
|
|
Title:
|
|
Crystal structure of apo-ll-diaminopimelate aminotransferase from arabidopsis thaliana (no plp)
|
|
Structure:
|
|
Ll-diaminopimelate aminotransferase. Chain: a, b. Fragment: unp residues 36 to 461. Synonym: ll-dap-aminotransferase, dap-aminotransferase, dap-at, atdap-at, protein aberrant growth and death 2. Engineered: yes
|
|
Source:
|
|
Arabidopsis thaliana. Mouse-ear cress,thale-cress. Organism_taxid: 3702. Gene: dap, agd2, at4g33680, t16l1.170. Expressed in: escherichia coli. Expression_system_taxid: 562
|
|
Resolution:
|
|
|
1.99Å
|
R-factor:
|
0.191
|
R-free:
|
0.229
|
|
|
Authors:
|
|
N.Watanabe,M.D.Clay,M.J.Van Belkum,M.M.Cherney,J.C.Vederas, M.N.G.James
|
Key ref:

|
|
N.Watanabe
et al.
(2008).
Mechanism of substrate recognition and PLP-induced conformational changes in LL-diaminopimelate aminotransferase from Arabidopsis thaliana.
J Mol Biol,
384,
1314-1329.
PubMed id:
DOI:

|
|
|
Date:
|
|
|
15-Sep-08
|
Release date:
|
14-Oct-08
|
|
|
|
|
|
|
PROCHECK
|

|
|
|
|
|
Headers
|
|
|
|
References
|
|
|
|
|
|

|
|
Q93ZN9
(DAPAT_ARATH) -
LL-diaminopimelate aminotransferase, chloroplastic from Arabidopsis thaliana
|
|
|
|
Seq:
Struc:
|
|
|
|
461 a.a.
412 a.a.
|
|

|

|

|
|
|
|
|
|
|
|
|
|
|
Key: |
 |
PfamA domain |
|
|
 |
Secondary structure |
|
 |
CATH domain |
|
|
|
|
|
|
|
|
|
|
|
|
|
Enzyme class:
|
|
E.C.2.6.1.83
- LL-diaminopimelate aminotransferase.
|
|
|
|
|
|
|
Reaction:
|
|
(2S,6S)-2,6-diaminopimelate + 2-oxoglutarate = (S)-2,3,4,5- tetrahydrodipicolinate + L-glutamate + H2O + H+
|
|
|
|
|
|
(2S,6S)-2,6-diaminopimelate
|
+
|
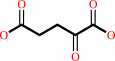 2-oxoglutarate
2-oxoglutarate
|
=
|
(S)-2,3,4,5- tetrahydrodipicolinate
|
+
|
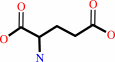 L-glutamate
L-glutamate
|
+
|
H2O
|
+
|
H(+)
|
|
|
|
|
|
|
|
|
|
Cofactor:
|
|
Pyridoxal 5'-phosphate
|
|
|
|
|
|
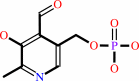 Pyridoxal 5'-phosphate
Pyridoxal 5'-phosphate
|
|
|
|
|
|
|
Molecule diagrams generated from .mol files obtained from the
KEGG ftp site
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
| |
|
|
| |
|
DOI no:
|
J Mol Biol
384:1314-1329
(2008)
|
|
PubMed id:
|
|
|
|
|
|
| |
|
Mechanism of substrate recognition and PLP-induced conformational changes in LL-diaminopimelate aminotransferase from Arabidopsis thaliana.
|
|
N.Watanabe,
M.D.Clay,
M.J.van Belkum,
M.M.Cherney,
J.C.Vederas,
M.N.James.
|
|
|
|
|
| |
ABSTRACT
|
|
|
|
| |
|
|
LL-Diaminopimelate aminotransferase (LL-DAP-AT), a pyridoxal phosphate
(PLP)-dependent enzyme in the lysine biosynthetic pathways of plants and
Chlamydia, is a potential target for the development of herbicides or
antibiotics. This homodimeric enzyme converts L-tetrahydrodipicolinic acid
(THDP) directly to LL-DAP using L-glutamate as the source of the amino group.
Earlier, we described the 3D structures of native and malate-bound LL-DAP-AT
from Arabidopsis thaliana (AtDAP-AT). Seven additional crystal structures of
AtDAP-AT and its variants are reported here as part of an investigation into the
mechanism of substrate recognition and catalysis. Two structures are of AtDAP-AT
with reduced external aldimine analogues: N-(5'-phosphopyridoxyl)-L-glutamate
(PLP-Glu) and N-(5'-phosphopyridoxyl)- LL-Diaminopimelate (PLP-DAP) bound in the
active site. Surprisingly, they reveal that both L-glutamate and LL-DAP are
recognized in a very similar fashion by the same sets of amino acid residues;
both molecules adopt twisted V-shaped conformations. With both substrates, the
alpha-carboxylates are bound in a salt bridge with Arg404, whereas the distal
carboxylates are recognized via hydrogen bonds to the well-conserved side chains
of Tyr37, Tyr125 and Lys129. The distal C(varepsilon) amino group of LL-DAP is
specifically recognized by several non-covalent interactions with residues from
the other subunit (Asn309*, Tyr94*, Gly95*, and Glu97* (Amino acid designators
followed by an asterisk (*) indicate that the residues originate in the other
subunit of the dimer)) and by three bound water molecules. Two catalytically
inactive variants of AtDAP-AT were created via site-directed mutagenesis of the
active site lysine (K270N and K270Q). The structures of these variants permitted
the observation of the unreduced external aldimines of PLP with L-glutamate and
with LL-DAP in the active site, and revealed differences in the torsion angle
about the PLP-substrate bond. Lastly, an apo-AtDAP-AT structure missing PLP
revealed details of conformational changes induced by PLP binding and substrate
entry into the active site.
|
|
|
|
|
|
| |
Selected figure(s)
|
|
|
|
| |
|
|

|
|

|
|
Figure 7.
Fig. 7. Two distinct conformations of Tyr364 in AtDAP-AT. (a)
An overlay of the crystal structures of the native AtDAP-AT and
the PLP-Glu aldimine analogue complex. The native structure is
coloured yellow. The PLP-Glu aldimine analogue structure is
coloured grey. Two different conformations of Tyr364 in the
active site are shown. (b) An overlay of aromatic residues
equivalent to Tyr364 of AtDAP-AT from the other type I fold
aminotransferases. AtDAP-AT, AspAT (Ia), AspAT (Ib) and HisP-AT
are represented in grey, yellow, blue and emerald green,
respectively.
|
|
Figure 8.
Fig. 8. An overlay of the structures of native AtDAP-AT,
K270N and K270Q variants of AtDAP-AT around active site Lys270
residue shown in line representation. The native, K270N and
K270Q variants are coloured cyan, green and magenta,
respectively.
|
|
|
|
|
|
| |
The above figures are
reprinted
by permission from Elsevier:
J Mol Biol
(2008,
384,
1314-1329)
copyright 2008.
|
|
| |
Figures were
selected
by an automated process.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
 |
 |
|
Literature references that cite this PDB file's key reference
|
|
|
| |
PubMed id
|
|
Reference
|
|
|
|
|
|
Y.Liu,
R.H.White,
and
W.B.Whitman
(2010).
Methanococci use the diaminopimelate aminotransferase (DapL) pathway for lysine biosynthesis.
|
| |
J Bacteriol,
192,
3304-3310.
|
|
|
|
|
|
The most recent references are shown first.
Citation data come partly from CiteXplore and partly
from an automated harvesting procedure. Note that this is likely to be
only a partial list as not all journals are covered by
either method. However, we are continually building up the citation data
so more and more references will be included with time.
|
|


 Links
Links




