 |
PDBsum entry 2qtg

|
|
|
|
|
 |
Contents |
 |
|
|
|
|
|
|
|
|
|
|
|
|
* Residue conservation analysis
|
|
|
|
|
|
|
|
|
|
|
Enzyme class:
|
|
E.C.3.2.2.16
- methylthioadenosine nucleosidase.
|
|
|
|
|
|
|
Reaction:
|
|
S-methyl-5'-thioadenosine + H2O = 5-(methylsulfanyl)-D-ribose + adenine
|
|
|
|
|
|
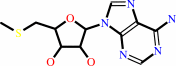 S-methyl-5'-thioadenosine
S-methyl-5'-thioadenosine
|
+
|
H2O
Bound ligand (Het Group name = )
matches with 90.48% similarity
|
=
|
5-(methylsulfanyl)-D-ribose
|
+
|
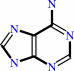 adenine
adenine
|
|
|
|
|
|
|
|
|
|
|
|
|
Molecule diagrams generated from .mol files obtained from the
KEGG ftp site
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
| |
|
|
| |
|
DOI no:
|
J Mol Biol
378:112-128
(2008)
|
|
PubMed id:
|
|
|
|
|
|
| |
|
Molecular determinants of substrate specificity in plant 5'-methylthioadenosine nucleosidases.
|
|
K.K.Siu,
J.E.Lee,
J.R.Sufrin,
B.A.Moffatt,
M.McMillan,
K.A.Cornell,
C.Isom,
P.L.Howell.
|
|
|
|
|
| |
ABSTRACT
|
|
|
|
| |
|
|
5'-Methylthioadenosine (MTA)/S-adenosylhomocysteine (SAH) nucleosidase (MTAN) is
essential for cellular metabolism and development in many bacterial species.
While the enzyme is found in plants, plant MTANs appear to select for MTA
preferentially, with little or no affinity for SAH. To understand what
determines substrate specificity in this enzyme, MTAN homologues from
Arabidopsis thaliana (AtMTAN1 and AtMTAN2, which are referred to as AtMTN1 and
AtMTN2 in the plant literature) have been characterized kinetically. While both
homologues hydrolyze MTA with comparable kinetic parameters, only AtMTAN2 shows
activity towards SAH. AtMTAN2 also has higher catalytic activity towards other
substrate analogues with longer 5'-substituents. The structures of apo AtMTAN1
and its complexes with the substrate- and transition-state-analogues,
5'-methylthiotubercidin and formycin A, respectively, have been determined at
2.0-1.8 A resolution. A homology model of AtMTAN2 was generated using the
AtMTAN1 structures. Comparison of the AtMTAN1 and AtMTAN2 structures reveals
that only three residues in the active site differ between the two enzymes. Our
analysis suggests that two of these residues, Leu181/Met168 and Phe148/Leu135 in
AtMTAN1/AtMTAN2, likely account for the divergence in specificity of the
enzymes. Comparison of the AtMTAN1 and available Escherichia coli MTAN (EcMTAN)
structures suggests that a combination of differences in the 5'-alkylthio
binding region and reduced conformational flexibility in the AtMTAN1 active site
likely contribute to its reduced efficiency in binding substrate analogues with
longer 5'-substituents. In addition, in contrast to EcMTAN, the active site of
AtMTAN1 remains solvated in its ligand-bound forms. As the apparent pK(a) of an
amino acid depends on its local environment, the putative catalytic acid Asp225
in AtMTAN1 may not be protonated at physiological pH and this suggests the
transition state of AtMTAN1, like human MTA phosphorylase and Streptococcus
pneumoniae MTAN, may be different from that found in EcMTAN.
|
|
|
|
|
|
| |
Selected figure(s)
|
|
|
|
| |
|
|

|
|

|
|
Figure 1.
Fig. 1. The structures of S-adenosylhomocysteine (SAH),
5′-methylthioadenosine (MTA), formycin A (FMA),
5′-methylthiotubercidin (MTT). For ease of comparison, the
structures are numbered according to MTA rather than the IUPAC
convention.
|
|
Figure 7.
Fig. 7. Conformational changes in the active sites of AtMTAN1
(a) and EcMTAN (b) upon binding of FMA or MTT. (c)
Superimposition of EcMTAN-Ade and apo-AtMTAN1. Electron density
is missing for the β10-α6 loop in EcMTAN because of disorder,
so this region is not modeled in the structure and is
represented here by a dotted line. (d) Superimposition of
EcMTAN-MTT and AtMTAN1-MTT highlighting the positions of Phe148
in AtMTAN1 and Tyr107 in EcMTAN and their relative proximities
to the 5′-alkylthio end of the ligand. In all panels, the apo-
and MTT-bound structures of AtMTAN1 are shown in yellow and
brown, respectively, while the ADE- and MTT-bound forms of
EcMTAN are shown in gray and pink, respectively.
|
|
|
|
|
|
| |
The above figures are
reprinted
by permission from Elsevier:
J Mol Biol
(2008,
378,
112-128)
copyright 2008.
|
|
| |
Figures were
selected
by an automated process.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
 |
 |
|
Literature references that cite this PDB file's key reference
|
|
|
| |
PubMed id
|
|
Reference
|
|
|
|
|
|
D.R.Ronning,
N.M.Iacopelli,
and
V.Mishra
(2010).
Enzyme-ligand interactions that drive active site rearrangements in the Helicobacter pylori 5'-methylthioadenosine/S-adenosylhomocysteine nucleosidase.
|
| |
Protein Sci,
19,
2498-2510.
|
|
|
PDB codes:
|
|
|
|
|
|
|
|
E.Albers
(2009).
Metabolic characteristics and importance of the universal methionine salvage pathway recycling methionine from 5'-methylthioadenosine.
|
| |
IUBMB Life,
61,
1132-1142.
|
|
|
|
|
|
The most recent references are shown first.
Citation data come partly from CiteXplore and partly
from an automated harvesting procedure. Note that this is likely to be
only a partial list as not all journals are covered by
either method. However, we are continually building up the citation data
so more and more references will be included with time.
Where a reference describes a PDB structure, the PDB
codes are
shown on the right.
|
|


 Links
Links











