 |
PDBsum entry 2ixi

|
|
|
|
|
 |
Contents |
 |
|
|
|
|
|
|
|
|
|
|
|
|
* Residue conservation analysis
|
|
|
|
|
|
|
|
|
|
|
Enzyme class:
|
|
E.C.5.1.3.13
- dTDP-4-dehydrorhamnose 3,5-epimerase.
|
|
|
|
|
|
|
Pathway:
|
|
6-Deoxyhexose Biosynthesis
|
|
|
|
|
|
Reaction:
|
|
dTDP-4-dehydro-6-deoxy-alpha-D-glucose = dTDP-4-dehydro-beta-L-rhamnose
|
|
|
|
|
|
dTDP-4-dehydro-6-deoxy-alpha-D-glucose
Bound ligand (Het Group name = )
matches with 71.43% similarity
|
=
|
dTDP-4-dehydro-beta-L-rhamnose
|
|
|
|
|
|
|
|
|
|
Cofactor:
|
|
NAD(+)
|
|
|
|
|
|
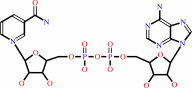 NAD(+)
NAD(+)
|
|
|
|
|
|
|
Molecule diagrams generated from .mol files obtained from the
KEGG ftp site
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
| |
|
|
| |
|
DOI no:
|
J Mol Biol
365:146-159
(2007)
|
|
PubMed id:
|
|
|
|
|
|
| |
|
RmlC, a C3' and C5' carbohydrate epimerase, appears to operate via an intermediate with an unusual twist boat conformation.
|
|
C.Dong,
L.L.Major,
V.Srikannathasan,
J.C.Errey,
M.F.Giraud,
J.S.Lam,
M.Graninger,
P.Messner,
M.R.McNeil,
R.A.Field,
C.Whitfield,
J.H.Naismith.
|
|
|
|
|
| |
ABSTRACT
|
|
|
|
| |
|
|
The striking feature of carbohydrates is their constitutional, conformational
and configurational diversity. Biology has harnessed this diversity and
manipulates carbohydrate residues in a variety of ways, one of which is
epimerization. RmlC catalyzes the epimerization of the C3' and C5' positions of
dTDP-6-deoxy-D-xylo-4-hexulose, forming dTDP-6-deoxy-L-lyxo-4-hexulose. RmlC is
the third enzyme of the rhamnose pathway, and represents a validated
anti-bacterial drug target. Although several structures of the enzyme have been
reported, the mechanism and the nature of the intermediates have remained
obscure. Despite its relatively small size (22 kDa), RmlC catalyzes four
stereospecific proton transfers and the substrate undergoes a major
conformational change during the course of the transformation. Here we report
the structure of RmlC from several organisms in complex with product and product
mimics. We have probed site-directed mutants by assay and by deuterium exchange.
The combination of structural and biochemical data has allowed us to assign key
residues and identify the conformation of the carbohydrate during turnover.
Clear knowledge of the chemical structure of RmlC reaction intermediates may
offer new opportunities for rational drug design.
|
|
|
|
|
|
| |
Selected figure(s)
|
|
|
|
| |
|
|

|
|

|
|
Figure 1.
Figure 1. (a) R = OdTDP. The RmlC reaction converts
dTDP-6-deoxy-D-xylo-4-hexulose to dTDP-6-deoxy-L-lyxo-4-hexulose
(thick arrow). This process involves a ring flip as well as
epimerization. The steps are shown according to the current
convention, however, this route goes through some very high
energy intermediates (notably the C1′, C3′, C5′ triaxial
product). (b) R = OGPD. The GME reaction converts GDP-D-mannose
to GDP-L-galactose. Shown boxed is the predicted twist boat
intermediate. The dominant order of the epimerization (C5′
first) and the ring flipped form of GDP-L-ribo-4-hexulose were
determined experimentally.^24 (c) R = OdTDP. Substrate and
product mimics employed in this study.
|
|
Figure 6.
Figure 6. (a) R = OdTDP. A possible mechanism for RmlC based
on structural and biochemical data. The key active site residues
are shown, the H65 is the catalytic base for both
epimerizations, K73 stabilizes the enolate and Y134 acts as the
acid for the first epimerization. The mono-epimerized
intermediate is shown boxed and has the equatorial linkage
between carbohydrate ring and nucleotide. It cannot proceed
directly to product because the C3′ proton is only
sufficiently acidic when it is orthogonal to the plane of the
carbonyl function. (b) R = OdTDP. An alternative route for the
first epimerization using a twist boat form of substrate, the
mono epimerized intermediate is shown boxed. The apparent
preference of RmlC for the equatorial linked sugar nucleotide
suggests that this is a possibility.
|
|
|
|
|
|
| |
The above figures are
reprinted
by permission from Elsevier:
J Mol Biol
(2007,
365,
146-159)
copyright 2007.
|
|
| |
Figures were
selected
by an automated process.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
 |
 |
|
Literature references that cite this PDB file's key reference
|
|
|
| |
PubMed id
|
|
Reference
|
|
|
|
|
|
L.M.Iyer,
S.Abhiman,
R.F.de Souza,
and
L.Aravind
(2010).
Origin and evolution of peptide-modifying dioxygenases and identification of the wybutosine hydroxylase/hydroperoxidase.
|
| |
Nucleic Acids Res,
38,
5261-5279.
|
|
|
|
|
|
|
S.Sivendran,
V.Jones,
D.Sun,
Y.Wang,
A.E.Grzegorzewicz,
M.S.Scherman,
A.D.Napper,
J.A.McCammon,
R.E.Lee,
S.L.Diamond,
and
M.McNeil
(2010).
Identification of triazinoindol-benzimidazolones as nanomolar inhibitors of the Mycobacterium tuberculosis enzyme TDP-6-deoxy-d-xylo-4-hexopyranosid-4-ulose 3,5-epimerase (RmlC).
|
| |
Bioorg Med Chem,
18,
896-908.
|
|
|
|
|
|
|
A.Caravano,
R.A.Field,
J.M.Percy,
G.Rinaudo,
R.Roig,
and
K.Singh
(2009).
Developing an asymmetric, stereodivergent route to selected 6-deoxy-6-fluoro-hexoses.
|
| |
Org Biomol Chem,
7,
996.
|
|
|
|
|
|
|
G.Agarwal,
M.Rajavel,
B.Gopal,
and
N.Srinivasan
(2009).
Structure-based phylogeny as a diagnostic for functional characterization of proteins with a cupin fold.
|
| |
PLoS One,
4,
e5736.
|
|
|
|
|
|
|
M.S.Byrd,
I.Sadovskaya,
E.Vinogradov,
H.Lu,
A.B.Sprinkle,
S.H.Richardson,
L.Ma,
B.Ralston,
M.R.Parsek,
E.M.Anderson,
J.S.Lam,
and
D.J.Wozniak
(2009).
Genetic and biochemical analyses of the Pseudomonas aeruginosa Psl exopolysaccharide reveal overlapping roles for polysaccharide synthesis enzymes in Psl and LPS production.
|
| |
Mol Microbiol,
73,
622-638.
|
|
|
|
|
|
|
R.A.Stabler,
M.He,
L.Dawson,
M.Martin,
E.Valiente,
C.Corton,
T.D.Lawley,
M.Sebaihia,
M.A.Quail,
G.Rose,
D.N.Gerding,
M.Gibert,
M.R.Popoff,
J.Parkhill,
G.Dougan,
and
B.W.Wren
(2009).
Comparative genome and phenotypic analysis of Clostridium difficile 027 strains provides insight into the evolution of a hypervirulent bacterium.
|
| |
Genome Biol,
10,
R102.
|
|
|
|
|
|
|
C.J.Thibodeaux,
C.E.Melançon,
and
H.W.Liu
(2008).
Natural-product sugar biosynthesis and enzymatic glycodiversification.
|
| |
Angew Chem Int Ed Engl,
47,
9814-9859.
|
|
|
|
|
|
|
M.Tello,
M.Rejzek,
B.Wilkinson,
D.M.Lawson,
and
R.A.Field
(2008).
Tyl1a, a TDP-6-deoxy-D-xylo-4-hexulose 3,4-isomerase from Streptomyces fradiae: structure prediction, mutagenesis and solvent isotope incorporation experiments to investigate reaction mechanism.
|
| |
Chembiochem,
9,
1295-1302.
|
|
|
|
|
|
|
J.D.King,
N.J.Harmer,
A.Preston,
C.M.Palmer,
M.Rejzek,
R.A.Field,
T.L.Blundell,
and
D.J.Maskell
(2007).
Predicting protein function from structure--the roles of short-chain dehydrogenase/reductase enzymes in Bordetella O-antigen biosynthesis.
|
| |
J Mol Biol,
374,
749-763.
|
|
|
PDB codes:
|
|
|
|
|
|
|
The most recent references are shown first.
Citation data come partly from CiteXplore and partly
from an automated harvesting procedure. Note that this is likely to be
only a partial list as not all journals are covered by
either method. However, we are continually building up the citation data
so more and more references will be included with time.
Where a reference describes a PDB structure, the PDB
codes are
shown on the right.
|
|


 Links
Links











