 |
PDBsum entry 1sd0

|
|
|
|
|
 |
Contents |
 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
* Residue conservation analysis
|
|
|
|
|
References listed in PDB file
|
|
|
Key reference
|
|
|
Title
|
|
The active site cysteine of arginine kinase: structural and functional analysis of partially active mutants.
|
|
|
Authors
|
|
J.L.Gattis,
E.Ruben,
M.O.Fenley,
W.R.Ellington,
M.S.Chapman.
|
|
|
Ref.
|
|
Biochemistry, 2004,
43,
8680-8689.
[DOI no: ]
|
|
|
PubMed id
|
|
|
|
|
|
Abstract
|
|
|
Arginine kinase buffers cellular ATP levels by catalyzing reversible phosphoryl
transfer between ATP and arginine. A conserved cysteine has long been thought
important in catalysis. Here, cysteine 271 of horseshoe crab arginine kinase has
been mutated to serine, alanine, asparagine, or aspartate. Catalytic turnover
rates were 0.02-1.0% of wild type, but the activity of uncharged mutations could
be partially rescued with chloride. Steady-state binding constants were slightly
increased, more so for phospho-L-arginine than ADP. Substrate binding synergy
observed in many phosphagen kinases was reduced or eliminated in mutant enzymes.
The crystallographic structure of the alanine mutant at 2.3 A resolution,
determined as a transition state analogue complex with arginine, nitrate, and
MgADP, was nearly identical to wild type. Enzyme-substrate interactions are
maintained as in wild type, and substrates remain at least roughly aligned for
in-line phosphoryl transfer. Homology models with serine, asparagine, or
aspartate replacing the active site cysteine similarly show only minor
structural changes. Most striking, however, is the presence in the C271A mutant
crystallographic structure of a chloride ion within 3.5 A of the nonreactive
N(eta) substrate nitrogen, approximating the position of the sulfur in the
wild-type's cysteine. Together, the results contradict prevailing speculation
that the cysteine mediates a substrate-induced conformational change, confirm
that it is the thiolate form that is relevant to catalysis, and suggest that one
of its roles is to help to enhance the catalytic rate through electrostatic
stabilization of the transition state.
|
|
|
Secondary reference #1
|
|
|
Title
|
|
Transition state structure of arginine kinase: implications for catalysis of bimolecular reactions.
|
|
|
Authors
|
|
G.Zhou,
T.Somasundaram,
E.Blanc,
G.Parthasarathy,
W.R.Ellington,
M.S.Chapman.
|
|
|
Ref.
|
|
Proc Natl Acad Sci U S A, 1998,
95,
8449-8454.
[DOI no: ]
|
|
|
PubMed id
|
|
|
|
|
|
|
|
|
|

|
|

|
|
Figure 2.
Fig. 2. Details of the active site. For clarity only
atoms in the immediate neighborhood are shown with
carbon-colored black, oxygen red, nitrogen dark blue, magnesium
light blue, sulfur yellow and phosphorus gray. Distances are
shown in Å. (a) Stereo diagram comparing part of the
experimental analog structure with omit-map electron density and
the structure of the presumptive transition state (gray atoms).
Small molecule model systems suggest a preassociative concerted
phosphoryl transfer and a pentavalent  -phosphorus
transition state with about 20% covalent bonding to both the -phosphorus
transition state with about 20% covalent bonding to both the
 -phosphoryl
oxygen and guanidino nitrogen (32, 33). The transition-state
coordinates were derived from the experimental coordinates by
replacing the nitrate with a phosphoryl group, and refining with
additional distance and angle restraints appropriate for the
estimated 20% partial covalent bonding. (b-e) Details of the
enzyme-substrate analog interactions: (b) -phosphoryl
oxygen and guanidino nitrogen (32, 33). The transition-state
coordinates were derived from the experimental coordinates by
replacing the nitrate with a phosphoryl group, and refining with
additional distance and angle restraints appropriate for the
estimated 20% partial covalent bonding. (b-e) Details of the
enzyme-substrate analog interactions: (b)  and phosphoryl
groups of the ADP are held in place by extensive hydrogen
bonds/salt bridges with four highly conserved arginines; (c) the
nitrate (mimicking a planar phosphoryl group during transfer) is
sandwiched between two conserved arginines and the Mg2+ ion
whose position is constrained by ligands from the and phosphoryl
groups of the ADP; (d) the guanidinium of the substrate arginine
is clamped with salt bridges/hydrogen bonds to two carboxylates
and a conserved cysteine that likely exists as a thiolate (54);
and (e) interactions of the substrate amino and carboxylate
groups with loop residues 63-68 of the enzyme. The
carboxylate-to-backbone interactions might be conserved between
all phosphagens and their kinases. The amino groups are present
in arginine and lombricine but absent from creatine and
glycocyamine. The tyrosine interacting with the amino group is
conserved among AKs, but is a valine in all other phosphagen
kinases. Immediately preceding residue 61 (and interactions with
the carboxylate) is an insertion in other sequences whose size
inversely correlates with the size of substrate (42). and phosphoryl
groups of the ADP are held in place by extensive hydrogen
bonds/salt bridges with four highly conserved arginines; (c) the
nitrate (mimicking a planar phosphoryl group during transfer) is
sandwiched between two conserved arginines and the Mg2+ ion
whose position is constrained by ligands from the and phosphoryl
groups of the ADP; (d) the guanidinium of the substrate arginine
is clamped with salt bridges/hydrogen bonds to two carboxylates
and a conserved cysteine that likely exists as a thiolate (54);
and (e) interactions of the substrate amino and carboxylate
groups with loop residues 63-68 of the enzyme. The
carboxylate-to-backbone interactions might be conserved between
all phosphagens and their kinases. The amino groups are present
in arginine and lombricine but absent from creatine and
glycocyamine. The tyrosine interacting with the amino group is
conserved among AKs, but is a valine in all other phosphagen
kinases. Immediately preceding residue 61 (and interactions with
the carboxylate) is an insertion in other sequences whose size
inversely correlates with the size of substrate (42).
|
|
Figure 3.
Fig. 3. Roles of neighboring amino acids in the catalytic
mechanism of AK: (a) the forward reaction and (b) the reverse
reaction. In this schematic representation, only the reactive
groups of participants are shown. *, The structure does not
indicate whether it is Glu-225 or Glu-314 that acts as the
proposed acid/base catalyst.
|
|
|
|
|
|
Secondary reference #2
|
|
|
Title
|
|
Refinement of the arginine kinase transition-State analogue complex at 1.2 a resolution: mechanistic insights.
|
|
|
Authors
|
|
M.S.Yousef,
F.Fabiola,
J.L.Gattis,
T.Somasundaram,
M.S.Chapman.
|
|
|
Ref.
|
|
Acta Crystallogr D Biol Crystallogr, 2002,
58,
2009-2017.
[DOI no: ]
|
|
|
PubMed id
|
|
|
|
|
|
|
|
|
|

|
|

|
|
Figure 2.
Figure 2 (a) Precise alignment of the substrates in the active
site as revealed by the 2F[o] - F[c] electron-density map
contoured at 3  .
(b) The in-line transfer of the phosphoryl group and the
hydrogen-bond geometry between O[ .
(b) The in-line transfer of the phosphoryl group and the
hydrogen-bond geometry between O[ 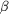 1]
of ADP and O[1] of nitrate. Angles in parentheses correspond to
the optimal reaction trajectory (Zhou et al., 1998[Zhou, G.,
Somasundaram, T., Blanc, E., Parthsarathy, G., Ellington, W. R.
& Chapman, M. S. (1998). Proc. Natl Acad. Sci. USA, 95,
8449-8454.]). 1]
of ADP and O[1] of nitrate. Angles in parentheses correspond to
the optimal reaction trajectory (Zhou et al., 1998[Zhou, G.,
Somasundaram, T., Blanc, E., Parthsarathy, G., Ellington, W. R.
& Chapman, M. S. (1998). Proc. Natl Acad. Sci. USA, 95,
8449-8454.]).
|
|
Figure 4.
Figure 4 Coordination of Mg2+ in the active site. Distances are
shown in �.
|
|
|
|
|
|
The above figures are
reproduced from the cited reference
with permission from the IUCr
|
|
|
Secondary reference #3
|
|
|
Title
|
|
Induced fit in guanidino kinases--Comparison of substrate-Free and transition state analog structures of arginine kinase.
|
|
|
Authors
|
|
M.S.Yousef,
S.A.Clark,
P.K.Pruett,
T.Somasundaram,
W.R.Ellington,
M.S.Chapman.
|
|
|
Ref.
|
|
Protein Sci, 2003,
12,
103-111.
[DOI no: ]
|
|
|
PubMed id
|
|
|
|
|
|
|
|
|
|

|
|

|
|
Figure 1.
Figure 1. Example electron density: a 2F[o] - F[c] omit
map, contoured at 1.5  , around
residues Phe[218], Leu[219], and Val[220]. , around
residues Phe[218], Leu[219], and Val[220].
|
|
Figure 3.
Figure 3. Space-filling models comparing open and closed
forms of arginine kinase. (A) The substrate-free conformation is
shown with the substrates in stick model as they would be bound
in the closed form. (B) The closed form shields the substrates
from solvent access.
|
|
|
|
|
|
The above figures are
reproduced from the cited reference
with permission from the Protein Society
|
|
|
|
|
|
|





