 |
PDBsum entry 1q2c

|
|
|
|
|
|

|

|

|

|
|

|
|

|
|

|
|

|
|
|
 |

|

|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Transferase/structural protein
|
PDB id
|
|
|
|
1q2c
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|

|

|

|
|
|
|
|
|
|
|
 |
Contents |
 |
|
|
|
|
|
|
|
|
|
|
|
|
* Residue conservation analysis
|
|
|
|
|
|
|
|
|
|
|
Enzyme class:
|
|
E.C.2.3.1.48
- histone acetyltransferase.
|
|
|
|
|
|
|
Reaction:
|
|
L-lysyl-[protein] + acetyl-CoA = N6-acetyl-L-lysyl-[protein] + CoA + H+
|
|
|
|
|
|
L-lysyl-[protein]
|
+
|
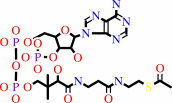 acetyl-CoA
acetyl-CoA
|
=
|
N(6)-acetyl-L-lysyl-[protein]
Bound ligand (Het Group name = )
matches with 95.92% similarity
|
+
|
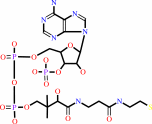 CoA
CoA
|
+
|
H(+)
|
|
|
|
|
|
|
|
|
|
|
|
|
Molecule diagrams generated from .mol files obtained from the
KEGG ftp site
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
| |
|
|
| |
|
DOI no:
|
Mol Cell
12:461-473
(2003)
|
|
PubMed id:
|
|
|
|
|
|
| |
|
Structural basis for histone and phosphohistone binding by the GCN5 histone acetyltransferase.
|
|
A.Clements,
A.N.Poux,
W.S.Lo,
L.Pillus,
S.L.Berger,
R.Marmorstein.
|
|
|
|
|
| |
ABSTRACT
|
|
|
|
| |
|
|
Distinct posttranslational modifications on histones occur in specific patterns
to mediate certain chromosomal events. For example, on histone H3,
phosphorylation at Ser10 can enhance GCN5-mediated Lys14 acetylation to promote
transcription. To gain insight into the mechanism underlying this synergism, we
determined the structure of Tetrahymena GCN5 (tGCN5) and coenzyme A (CoA) bound
to unmodified and Ser10-phosphorylated 19 residue histone H3 peptides (H3p19 and
H3p19Pi, respectively). The tGCN5/CoA/H3p19 structure reveals that a 12 amino
acid core sequence mediates extensive contacts with the protein, providing the
structural basis for substrate specificity by the GCN5/PCAF family of histone
acetyltransferases. Comparison with the tGCN5/CoA/H3p19Pi structure reveals that
phospho-Ser10 and Thr11 mediate significant histone-protein interactions, and
nucleate additional interactions distal to the phosphorylation site. Functional
studies show that histone H3 Thr11 is necessary for optimal transcription at
yGcn5-dependent promoters requiring Ser10 phosphorylation. Together, these
studies reveal how one histone modification can modulate another to affect
distinct transcriptional signals.
|
|
|
|
|
|
| |
Selected figure(s)
|
|
|
|
| |
|
|

|
|

|
|
Figure 3.
Figure 3. Comparison between the tGCN5/CoA/H3p19 and
tGCN5/CoA/H3p19Pi Structures(A) The H3p19Pi peptide N termini.
The tGCN5 blue residues represent amino acids that contact the
same residues in both peptides. The tGCN5 aqua residues
represent amino acids that are contacting additional residues in
H3p19Pi. The tGCN5 dark green residues represent amino acids
that interact with H3p19 and not with any residues in H3p19Pi.
For simplicity, hydrogen-bonding interactions between H3p19Pi
and tGCN5 are not shown in this figure. The residue shown in
purple is a tGCN5 arginine from a symmetry-related molecule that
is interacting with tGCN5 in close proximity with the
peptide.(B) The H3p19Pi-peptide C termini. Hydrogen bonding
between H3p19Pi and tGCN5 is represented by red dashed lines.
Waters are represented as yellow spheres.(C and D) Hydrogen
bonding interactions between phospho-Ser10 and tGCN5 in both
conformations. The tGCN5 blue residues represent amino acids
that contact Ser10 of H3p19Pi, and hydrogen bonding is
represented by dashed lines.(E) Summary of the H3p19 and
H3p19Pi-derived peptide-tGCN5 interactions. The gray regions of
the peptide represent side chains/residues modeled in the
H3p19Pi peptide and not modeled for the H3p19 peptide. The
phosphate of S10P is in red. The bold bonds are shown for
residues that have an increased buried surface area of at least
50 Å^2 with tGCN5/CoA when compared to the tGCN5/CoA/H3p19
interactions. The solid and dashed arrows represent tGCN5
residues that are within H-bonding and van der Waals packing
distance of the peptide, respectively. Residues in blue
represent amino acids that contact the same residues in both
peptides. Residues in aqua represent amino acids that are
contacting residues only in H3p19Pi. The tGCN5 brown residues
represent amino acids that only contact S10PA. Residues in green
interact with H3p19 only. The tGCN5 salmon-colored residues
represent amino acids that only contact S10PB. Symbols
representing residue conservation and mutational sensitivity are
the same as described in the legend to Figure 1E.
|
|
Figure 6.
Figure 6. Sequence Comparisons of GCN5/PCAF
Substrates^aSubstrate specificity ratios are calculated relative
to H3p19^35. Substrate lysines for the sequences are identified
by: ^bGrant et al., 1999; ^cKuo et al., 1996; ^dLiu et al. 1999
and Sakaguchi et al. 1998; ^eMunshi et al., 1998; ^fSartorelli
et al., 1999; ^gKiernan et al., 1999.
|
|
|
|
|
|
| |
The above figures are
reprinted
by permission from Cell Press:
Mol Cell
(2003,
12,
461-473)
copyright 2003.
|
|
| |
Figures were
selected
by an automated process.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
 |
 |
|
Literature references that cite this PDB file's key reference
|
|
|
| |
PubMed id
|
|
Reference
|
|
|
|
|
|
S.Liokatis,
A.Stützer,
S.J.Elsässer,
F.X.Theillet,
R.Klingberg,
B.van Rossum,
D.Schwarzer,
C.D.Allis,
W.Fischle,
and
P.Selenko
(2012).
Phosphorylation of histone H3 Ser10 establishes a hierarchy for subsequent intramolecular modification events.
|
| |
Nat Struct Mol Biol,
19,
819-823.
|
|
|
|
|
|
|
R.F.Thompson,
M.J.Fazzari,
and
J.M.Greally
(2010).
Experimental approaches to the study of epigenomic dysregulation in ageing.
|
| |
Exp Gerontol,
45,
255-268.
|
|
|
|
|
|
|
B.Pérez-Cadahía,
B.Drobic,
and
J.R.Davie
(2009).
H3 phosphorylation: dual role in mitosis and interphase.
|
| |
Biochem Cell Biol,
87,
695-709.
|
|
|
|
|
|
|
C.Dinant,
A.B.Houtsmuller,
and
W.Vermeulen
(2008).
Chromatin structure and DNA damage repair.
|
| |
Epigenetics Chromatin,
1,
9.
|
|
|
|
|
|
|
C.E.Berndsen,
and
J.M.Denu
(2008).
Catalysis and substrate selection by histone/protein lysine acetyltransferases.
|
| |
Curr Opin Struct Biol,
18,
682-689.
|
|
|
|
|
|
|
I.E.Sánchez,
P.Beltrao,
F.Stricher,
J.Schymkowitz,
J.Ferkinghoff-Borg,
F.Rousseau,
and
L.Serrano
(2008).
Genome-wide prediction of SH2 domain targets using structural information and the FoldX algorithm.
|
| |
PLoS Comput Biol,
4,
e1000052.
|
|
|
|
|
|
|
K.D.Meyer,
A.J.Donner,
M.T.Knuesel,
A.G.York,
J.M.Espinosa,
and
D.J.Taatjes
(2008).
Cooperative activity of cdk8 and GCN5L within Mediator directs tandem phosphoacetylation of histone H3.
|
| |
EMBO J,
27,
1447-1457.
|
|
|
|
|
|
|
M.Shimada,
H.Niida,
D.H.Zineldeen,
H.Tagami,
M.Tanaka,
H.Saito,
and
M.Nakanishi
(2008).
Chk1 is a histone H3 threonine 11 kinase that regulates DNA damage-induced transcriptional repression.
|
| |
Cell,
132,
221-232.
|
|
|
|
|
|
|
W.Walter,
D.Clynes,
Y.Tang,
R.Marmorstein,
J.Mellor,
and
S.L.Berger
(2008).
14-3-3 interaction with histone H3 involves a dual modification pattern of phosphoacetylation.
|
| |
Mol Cell Biol,
28,
2840-2849.
|
|
|
|
|
|
|
Z.A.Gurard-Levin,
and
M.Mrksich
(2008).
The activity of HDAC8 depends on local and distal sequences of its peptide substrates.
|
| |
Biochemistry,
47,
6242-6250.
|
|
|
|
|
|
|
A.Schuetz,
G.Bernstein,
A.Dong,
T.Antoshenko,
H.Wu,
P.Loppnau,
A.Bochkarev,
and
A.N.Plotnikov
(2007).
Crystal structure of a binary complex between human GCN5 histone acetyltransferase domain and acetyl coenzyme A.
|
| |
Proteins,
68,
403-407.
|
|
|
PDB code:
|
|
|
|
|
|
|
|
C.E.Berndsen,
W.Selleck,
S.J.McBryant,
J.C.Hansen,
S.Tan,
and
J.M.Denu
(2007).
Nucleosome recognition by the Piccolo NuA4 histone acetyltransferase complex.
|
| |
Biochemistry,
46,
2091-2099.
|
|
|
|
|
|
|
K.K.Lee,
and
J.L.Workman
(2007).
Histone acetyltransferase complexes: one size doesn't fit all.
|
| |
Nat Rev Mol Cell Biol,
8,
284-295.
|
|
|
|
|
|
|
M.A.Holbert,
T.Sikorski,
J.Carten,
D.Snowflack,
S.Hodawadekar,
and
R.Marmorstein
(2007).
The human monocytic leukemia zinc finger histone acetyltransferase domain contains DNA-binding activity implicated in chromatin targeting.
|
| |
J Biol Chem,
282,
36603-36613.
|
|
|
PDB code:
|
|
|
|
|
|
|
|
S.A.Morris,
B.Rao,
B.A.Garcia,
S.B.Hake,
R.L.Diaz,
J.Shabanowitz,
D.F.Hunt,
C.D.Allis,
J.D.Lieb,
and
B.D.Strahl
(2007).
Identification of histone H3 lysine 36 acetylation as a highly conserved histone modification.
|
| |
J Biol Chem,
282,
7632-7640.
|
|
|
|
|
|
|
S.Lall
(2007).
Primers on chromatin.
|
| |
Nat Struct Mol Biol,
14,
1110-1115.
|
|
|
|
|
|
|
S.P.Baker,
and
P.A.Grant
(2007).
The SAGA continues: expanding the cellular role of a transcriptional co-activator complex.
|
| |
Oncogene,
26,
5329-5340.
|
|
|
|
|
|
|
T.Kouzarides
(2007).
Chromatin modifications and their function.
|
| |
Cell,
128,
693-705.
|
|
|
|
|
|
|
X.Dong,
M.Kato-Murayama,
T.Muramatsu,
H.Mori,
M.Shirouzu,
Y.Bessho,
and
S.Yokoyama
(2007).
The crystal structure of leucyl/phenylalanyl-tRNA-protein transferase from Escherichia coli.
|
| |
Protein Sci,
16,
528-534.
|
|
|
PDB code:
|
|
|
|
|
|
|
|
J.F.Couture,
and
R.C.Trievel
(2006).
Histone-modifying enzymes: encrypting an enigmatic epigenetic code.
|
| |
Curr Opin Struct Biol,
16,
753-760.
|
|
|
|
|
|
|
S.H.Ahn,
R.L.Diaz,
M.Grunstein,
and
C.D.Allis
(2006).
Histone H2B deacetylation at lysine 11 is required for yeast apoptosis induced by phosphorylation of H2B at serine 10.
|
| |
Mol Cell,
24,
211-220.
|
|
|
|
|
|
|
Y.Zhang,
B.Gilquin,
S.Khochbin,
and
P.Matthias
(2006).
Two catalytic domains are required for protein deacetylation.
|
| |
J Biol Chem,
281,
2401-2404.
|
|
|
|
|
|
|
Z.Ge,
C.Liu,
M.Björkholm,
A.Gruber,
and
D.Xu
(2006).
Mitogen-activated protein kinase cascade-mediated histone H3 phosphorylation is critical for telomerase reverse transcriptase expression/telomerase activation induced by proliferation.
|
| |
Mol Cell Biol,
26,
230-237.
|
|
|
|
|
|
|
K.Ingvarsdottir,
N.J.Krogan,
N.C.Emre,
A.Wyce,
N.J.Thompson,
A.Emili,
T.R.Hughes,
J.F.Greenblatt,
and
S.L.Berger
(2005).
H2B ubiquitin protease Ubp8 and Sgf11 constitute a discrete functional module within the Saccharomyces cerevisiae SAGA complex.
|
| |
Mol Cell Biol,
25,
1162-1172.
|
|
|
|
|
|
|
M.Biel,
V.Wascholowski,
and
A.Giannis
(2005).
Epigenetics--an epicenter of gene regulation: histones and histone-modifying enzymes.
|
| |
Angew Chem Int Ed Engl,
44,
3186-3216.
|
|
|
|
|
|
|
M.K.Shirra,
S.E.Rogers,
D.E.Alexander,
and
K.M.Arndt
(2005).
The Snf1 protein kinase and Sit4 protein phosphatase have opposing functions in regulating TATA-binding protein association with the Saccharomyces cerevisiae INO1 promoter.
|
| |
Genetics,
169,
1957-1972.
|
|
|
|
|
|
|
N.Macdonald,
J.P.Welburn,
M.E.Noble,
A.Nguyen,
M.B.Yaffe,
D.Clynes,
J.G.Moggs,
G.Orphanides,
S.Thomson,
J.W.Edmunds,
A.L.Clayton,
J.A.Endicott,
and
L.C.Mahadevan
(2005).
Molecular basis for the recognition of phosphorylated and phosphoacetylated histone h3 by 14-3-3.
|
| |
Mol Cell,
20,
199-211.
|
|
|
PDB codes:
|
|
|
|
|
|
|
|
P.O.Hassa,
and
M.O.Hottiger
(2005).
An epigenetic code for DNA damage repair pathways?
|
| |
Biochem Cell Biol,
83,
270-285.
|
|
|
|
|
|
|
R.T.Utley,
N.Lacoste,
O.Jobin-Robitaille,
S.Allard,
and
J.Côté
(2005).
Regulation of NuA4 histone acetyltransferase activity in transcription and DNA repair by phosphorylation of histone H4.
|
| |
Mol Cell Biol,
25,
8179-8190.
|
|
|
|
|
|
|
S.Huang,
M.Litt,
and
G.Felsenfeld
(2005).
Methylation of histone H4 by arginine methyltransferase PRMT1 is essential in vivo for many subsequent histone modifications.
|
| |
Genes Dev,
19,
1885-1893.
|
|
|
|
|
|
|
W.S.Lo,
E.R.Gamache,
K.W.Henry,
D.Yang,
L.Pillus,
and
S.L.Berger
(2005).
Histone H3 phosphorylation can promote TBP recruitment through distinct promoter-specific mechanisms.
|
| |
EMBO J,
24,
997.
|
|
|
|
|
|
|
Y.Liu,
X.Xu,
S.Singh-Rodriguez,
Y.Zhao,
and
M.H.Kuo
(2005).
Histone H3 Ser10 phosphorylation-independent function of Snf1 and Reg1 proteins rescues a gcn5- mutant in HIS3 expression.
|
| |
Mol Cell Biol,
25,
10566-10579.
|
|
|
|
|
|
|
C.J.Fry,
M.A.Shogren-Knaak,
and
C.L.Peterson
(2004).
Histone H3 amino-terminal tail phosphorylation and acetylation: synergistic or independent transcriptional regulatory marks?
|
| |
Cold Spring Harb Symp Quant Biol,
69,
219-226.
|
|
|
|
|
|
|
L.F.Chen,
and
W.C.Greene
(2004).
Shaping the nuclear action of NF-kappaB.
|
| |
Nat Rev Mol Cell Biol,
5,
392-401.
|
|
|
|
|
|
|
L.Johnson,
S.Mollah,
B.A.Garcia,
T.L.Muratore,
J.Shabanowitz,
D.F.Hunt,
and
S.E.Jacobsen
(2004).
Mass spectrometry analysis of Arabidopsis histone H3 reveals distinct combinations of post-translational modifications.
|
| |
Nucleic Acids Res,
32,
6511-6518.
|
|
|
|
|
|
|
M.J.Bottomley
(2004).
Structures of protein domains that create or recognize histone modifications.
|
| |
EMBO Rep,
5,
464-469.
|
|
|
|
|
|
|
N.C.Emre,
and
S.L.Berger
(2004).
Histone H2B ubiquitylation and deubiquitylation in genomic regulation.
|
| |
Cold Spring Harb Symp Quant Biol,
69,
289-299.
|
|
|
|
|
|
|
A.N.Poux,
and
R.Marmorstein
(2003).
Molecular basis for Gcn5/PCAF histone acetyltransferase selectivity for histone and nonhistone substrates.
|
| |
Biochemistry,
42,
14366-14374.
|
|
|
PDB code:
|
|
|
|
|
|
|
The most recent references are shown first.
Citation data come partly from CiteXplore and partly
from an automated harvesting procedure. Note that this is likely to be
only a partial list as not all journals are covered by
either method. However, we are continually building up the citation data
so more and more references will be included with time.
Where a reference describes a PDB structure, the PDB
code is
shown on the right.
|
|


 Links
Links













