 |
PDBsum entry 1mss

|
|
|
|
|
|

|

|

|

|
|

|
|

|
|

|
|

|
|

|
|
|
 |

|

|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Isomerase(intramolecular oxidoreductase)
|
PDB id
|
|
|
|
1mss
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|

|

|

|
|
|
|
|
|
|
|
 |
Contents |
 |
|
|
|
|
|
|
|
|
|
|
* Residue conservation analysis
|
|
|
|
|
|
|
|
|
|
|
Enzyme class:
|
|
E.C.5.3.1.1
- triose-phosphate isomerase.
|
|
|
|
|
|
|
Reaction:
|
|
D-glyceraldehyde 3-phosphate = dihydroxyacetone phosphate
|
|
|
|
|
|
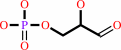 D-glyceraldehyde 3-phosphate
D-glyceraldehyde 3-phosphate
|
=
|
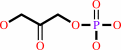 dihydroxyacetone phosphate
dihydroxyacetone phosphate
|
|
|
|
|
|
|
|
|
|
|
|
|
Molecule diagrams generated from .mol files obtained from the
KEGG ftp site
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
| |
|
|
| |
|
DOI no:
|
Structure
3:669-679
(1995)
|
|
PubMed id:
|
|
|
|
|
|
| |
|
Three new crystal structures of point mutation variants of monoTIM: conformational flexibility of loop-1, loop-4 and loop-8.
|
|
T.V.Borchert,
K.V.Kishan,
J.P.Zeelen,
W.Schliebs,
N.Thanki,
R.Abagyan,
R.Jaenicke,
R.K.Wierenga.
|
|
|
|
|
| |
ABSTRACT
|
|
|
|
| |
|
|
BACKGROUND: Wild-type triosephosphate isomerase (TIM) is a very stable dimeric
enzyme. This dimer can be converted into a stable monomeric protein (monoTIM) by
replacing the 15-residue interface loop (loop-3) by a shorter, 8-residue, loop.
The crystal structure of monoTIM shows that two active-site loops (loop-1 and
loop-4), which are at the dimer interface in wild-type TIM, have acquired rather
different structural properties. Nevertheless, monoTIM has residual catalytic
activity. RESULTS: Three new structures of variants of monoTIM are presented, a
double-point mutant crystallized in the presence and absence of bound inhibitor,
and a single-point mutant in the presence of a different inhibitor. These new
structures show large structural variability for the active-site loops, loop-1,
loop-4 and loop-8. In the structures with inhibitor bound, the catalytic lysine
(Lys13 in loop-1) and the catalytic histidine (His95 in loop-4) adopt
conformations similar to those observed in wild-type TIM, but very different
from the monoTIM structure. CONCLUSIONS: The residual catalytic activity of
monoTIM can now be rationalized. In the presence of substrate analogues the
active-site loops, loop-1, loop-4 and loop-8, as well as the catalytic residues,
adopt conformations similar to those seen in the wild-type protein. These loops
lack conformational flexibility in wild-type TIM. The data suggest that the
rigidity of these loops in wild-type TIM, resulting from subunit-subunit
contacts at the dimer interface, is important for optimal catalysis.
|
|
|
|
|
|
| |
Selected figure(s)
|
|
|
|
| |
|
|

|
|

|
|
Figure 1.
Figure 1. (a) The reaction catalyzed by TIM. (b) The structures
of two inhibitors of TIM, PGH and 2PG. Figure 1. (a) The
reaction catalyzed by TIM. (b) The structures of two inhibitors
of TIM, PGH and 2PG.
|
|
Figure 4.
Figure 4. Comparison of monoTIM-SS (red), monoTIM (green) and
wtTIM (yellow). (a) The complete Cα traces. One residue of each
front loop of monoTIM is labelled: Trp12 (L1), Phe45 (L2),
Ala70 (L3), His95 (L4), Glu129 (L5), Val169 (L6), Gly212 (L7)
and Gly235 (L8). Also shown, in green, is the sulphate ion,
near loop-6, loop-7 and loop-8, as observed in the monoTIM
structure. (b) Comparison of the Cα traces near L8, L1, L2 and
L3. The side chains of residues Ser237 (L8), Trp12 (L1), Thr44
(L2) and Gln65 (L3) are also shown. Figure 4. Comparison of
monoTIM-SS (red), monoTIM (green) and wtTIM (yellow). (a) The
complete Cα traces. One residue of each front loop of monoTIM
is labelled: Trp12 (L1), Phe45 (L2), Ala70 (L3), His95 (L4),
Glu129 (L5), Val169 (L6), Gly212 (L7) and Gly235 (L8). Also
shown, in green, is the sulphate ion, near loop-6, loop-7 and
loop-8, as observed in the monoTIM structure. (b) Comparison of
the Cα traces near L8, L1, L2 and L3. The side chains of
residues Ser237 (L8), Trp12 (L1), Thr44 (L2) and Gln65 (L3) are
also shown.
|
|
|
|
|
|
| |
The above figures are
reprinted
by permission from Cell Press:
Structure
(1995,
3,
669-679)
copyright 1995.
|
|
| |
Figures were
selected
by an automated process.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
 |
 |
|
Literature references that cite this PDB file's key reference
|
|
|
| |
PubMed id
|
|
Reference
|
|
|
|
|
|
M.Salin,
E.G.Kapetaniou,
M.Vaismaa,
M.Lajunen,
M.G.Casteleijn,
P.Neubauer,
L.Salmon,
and
R.K.Wierenga
(2010).
Crystallographic binding studies with an engineered monomeric variant of triosephosphate isomerase.
|
| |
Acta Crystallogr D Biol Crystallogr,
66,
934-944.
|
|
|
PDB codes:
|
|
|
|
|
|
|
|
R.K.Wierenga,
E.G.Kapetaniou,
and
R.Venkatesan
(2010).
Triosephosphate isomerase: a highly evolved biocatalyst.
|
| |
Cell Mol Life Sci,
67,
3961-3982.
|
|
|
|
|
|
|
M.Alahuhta,
M.G.Casteleijn,
P.Neubauer,
and
R.K.Wierenga
(2008).
Structural studies show that the A178L mutation in the C-terminal hinge of the catalytic loop-6 of triosephosphate isomerase (TIM) induces a closed-like conformation in dimeric and monomeric TIM.
|
| |
Acta Crystallogr D Biol Crystallogr,
64,
178-188.
|
|
|
PDB codes:
|
|
|
|
|
|
|
|
T.Cardozo,
T.Kimura,
S.Philpott,
B.Weiser,
H.Burger,
and
S.Zolla-Pazner
(2007).
Structural basis for coreceptor selectivity by the HIV type 1 V3 loop.
|
| |
AIDS Res Hum Retroviruses,
23,
415-426.
|
|
|
|
|
|
|
T.Prakash,
K.S.Sandhu,
N.K.Singh,
Y.Bhasin,
C.Ramakrishnan,
and
S.K.Brahmachari
(2007).
Structural assessment of glycyl mutations in invariantly conserved motifs.
|
| |
Proteins,
69,
617-632.
|
|
|
|
|
|
|
B.V.Norledge,
A.M.Lambeir,
R.A.Abagyan,
A.Rottmann,
A.M.Fernandez,
V.V.Filimonov,
M.G.Peter,
and
R.K.Wierenga
(2001).
Modeling, mutagenesis, and structural studies on the fully conserved phosphate-binding loop (loop 8) of triosephosphate isomerase: toward a new substrate specificity.
|
| |
Proteins,
42,
383-389.
|
|
|
PDB code:
|
|
|
|
|
|
|
|
A.M.Lambeir,
J.Backmann,
J.Ruiz-Sanz,
V.Filimonov,
J.E.Nielsen,
I.Kursula,
B.V.Norledge,
and
R.K.Wierenga
(2000).
The ionization of a buried glutamic acid is thermodynamically linked to the stability of Leishmania mexicana triose phosphate isomerase.
|
| |
Eur J Biochem,
267,
2516-2524.
|
|
|
PDB code:
|
|
|
|
|
|
|
|
R.Thoma,
M.Hennig,
R.Sterner,
and
K.Kirschner
(2000).
Structure and function of mutationally generated monomers of dimeric phosphoribosylanthranilate isomerase from Thermotoga maritima.
|
| |
Structure,
8,
265-276.
|
|
|
PDB code:
|
|
|
|
|
|
|
|
S.Miller,
B.Schuler,
and
R.Seckler
(1998).
A reversibly unfolding fragment of P22 tailspike protein with native structure: the isolated beta-helix domain.
|
| |
Biochemistry,
37,
9160-9168.
|
|
|
|
|
|
|
W.Schliebs,
N.Thanki,
R.Eritja,
and
R.Wierenga
(1996).
Active site properties of monomeric triosephosphate isomerase (monoTIM) as deduced from mutational and structural studies.
|
| |
Protein Sci,
5,
229-239.
|
|
|
|
|
|
The most recent references are shown first.
Citation data come partly from CiteXplore and partly
from an automated harvesting procedure. Note that this is likely to be
only a partial list as not all journals are covered by
either method. However, we are continually building up the citation data
so more and more references will be included with time.
Where a reference describes a PDB structure, the PDB
codes are
shown on the right.
|
|


 Links
Links











