 |
PDBsum entry 1fwt

|
|
|
|
|
 |
Contents |
 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
* Residue conservation analysis
|
|
|
|
|
|
PDB id:
|
|
|
|
| Name: |
|
Lyase
|
|
|
Title:
|
|
Aquifex aeolicus kdo8p synthase in complex with pep, e4p and cadmium
|
|
Structure:
|
|
2-dehydro-3-deoxyphosphooctonate aldolase. Chain: a, b. Synonym: kdo8p synthase, phospho-2-dehydro-3-deoxyoctonate aldolase, 3-deoxy-d-manno-octulosonic acid 8-phosphate synthetase, kdo-8- phosphate synthetase. Engineered: yes
|
|
Source:
|
|
Aquifex aeolicus. Organism_taxid: 63363. Expressed in: escherichia coli. Expression_system_taxid: 562.
|
|
Biol. unit:
|
|
 Tetramer (from PDB file)
Tetramer (from PDB file)
|
|
Resolution:
|
|
|
1.90Å
|
R-factor:
|
0.197
|
R-free:
|
0.222
|
|
|
Authors:
|
|
H.S.Duewel,S.Radaev,J.Wang,R.W.Woodard,D.L.Gatti
|
Key ref:

|
|
H.S.Duewel
et al.
(2001).
Substrate and metal complexes of 3-deoxy-D-manno-octulosonate-8-phosphate synthase from Aquifex aeolicus at 1.9-A resolution. Implications for the condensation mechanism.
J Biol Chem,
276,
8393-8402.
PubMed id:
DOI:

|
|
|
Date:
|
|
|
24-Sep-00
|
Release date:
|
21-Apr-01
|
|
|
|
|
|
|
PROCHECK
|

|
|
|
|
|
Headers
|
|
|
|
References
|
|
|
|
|
|

|
|
O66496
(KDSA_AQUAE) -
2-dehydro-3-deoxyphosphooctonate aldolase from Aquifex aeolicus (strain VF5)
|
|
|
|
Seq:
Struc:
|
|
|
|
267 a.a.
263 a.a.
|
|

|

|
|
|
|
|
|
|
|
|
|
|
|
Key: |
 |
PfamA domain |
|
|
 |
Secondary structure |
|
 |
CATH domain |
|
|
|
|
|
|
|
|
|
|
|
|
|
Enzyme class:
|
|
E.C.2.5.1.55
- 3-deoxy-8-phosphooctulonate synthase.
|
|
|
|
|
|
|
Reaction:
|
|
D-arabinose 5-phosphate + phosphoenolpyruvate + H2O = 3-deoxy-alpha-D- manno-2-octulosonate-8-phosphate + phosphate
|
|
|
|
|
|
D-arabinose 5-phosphate
Bound ligand (Het Group name = )
matches with 85.71% similarity
|
+
|
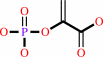 phosphoenolpyruvate
phosphoenolpyruvate
|
+
|
H2O
Bound ligand (Het Group name = )
corresponds exactly
|
=
|
3-deoxy-alpha-D- manno-2-octulosonate-8-phosphate
|
+
|
 phosphate
phosphate
|
|
|
|
|
|
|
|
|
|
|
|
|
Molecule diagrams generated from .mol files obtained from the
KEGG ftp site
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
| |
|
|
| |
|
DOI no:
|
J Biol Chem
276:8393-8402
(2001)
|
|
PubMed id:
|
|
|
|
|
|
| |
|
Substrate and metal complexes of 3-deoxy-D-manno-octulosonate-8-phosphate synthase from Aquifex aeolicus at 1.9-A resolution. Implications for the condensation mechanism.
|
|
H.S.Duewel,
S.Radaev,
J.Wang,
R.W.Woodard,
D.L.Gatti.
|
|
|
|
|
| |
ABSTRACT
|
|
|
|
| |
|
|
3-Deoxy-D-manno-octulosonate-8-phosphate synthase (KDO8PS) from the
hyperthermophilic bacterium Aquifex aeolicus differs from its Escherichia coli
counterpart in the requirement of a divalent metal for activity (Duewel, H. S.,
and Woodard, R. W. (2000) J. Biol. Chem. 275, 22824-22831). Here we report the
crystal structure of the A. aeolicus enzyme, which was determined by molecular
replacement using E. coli KDO8PS as a model. The structures of the metal-free
and Cd(2+) forms of the enzyme were determined in the uncomplexed state and in
complex with various combinations of phosphoenolpyruvate (PEP), arabinose
5-phosphate (A5P), and erythrose 4-phosphate (E4P). Like the E. coli enzyme, A.
aeolicus KDO8PS is a homotetramer containing four distinct active sites at the
interface between subunits. The active site cavity is open in the substrate-free
enzyme or when either A5P alone or PEP alone binds, and becomes isolated from
the aqueous phase when both PEP and A5P (or E4P) bind together. In the presence
of metal, the enzyme is asymmetric and appears to alternate catalysis between
the active sites located on one face of the tetramer and those located on the
other face. In the absence of metal, the asymmetry is lost. Details of the
active site that may be important for catalysis are visible at the high
resolution achieved in these structures. Most notably, the shape of the
PEP-binding pocket forces PEP to assume a distorted geometry at C-2, which might
anticipate the conversion from sp(2) to sp(3) hybridization occurring during
intermediate formation and which may modulate PEP reactivity toward A5P. Two
water molecules are located in van der Waals contact with the si and re sides of
C-2(PEP), respectively. Abstraction of a proton from either of these water
molecules by a protein group is expected to elicit a nucleophilic attack of the
resulting hydroxide ion on the nearby C-2(PEP), thus triggering the beginning of
the catalytic cycle.
|
|
|
|
|
|
| |
Selected figure(s)
|
|
|
|
| |
|
|

|
|

|
|
Figure 3.
Fig. 3. Metal-binding site of A. aeolicus KDO8PS. The C-
 trace and
side chains of the Cd^2+ enzyme are shown with salmon and white
bonds, respectively. The C- trace and
side chains of the metal-free enzyme are shown with transparent
light-blue bonds. The coordination of the Cd^2+ ion is shown as
transparent light-green bonds. Phosphate ions are labeled
PO[4]-1 and PO[4]-2. Blue, nitrogen; red, oxygen; yellow,
sulfur; pale blue, phosphorus; cyan, cadmium. WAT, water. trace and
side chains of the Cd^2+ enzyme are shown with salmon and white
bonds, respectively. The C- trace and
side chains of the metal-free enzyme are shown with transparent
light-blue bonds. The coordination of the Cd^2+ ion is shown as
transparent light-green bonds. Phosphate ions are labeled
PO[4]-1 and PO[4]-2. Blue, nitrogen; red, oxygen; yellow,
sulfur; pale blue, phosphorus; cyan, cadmium. WAT, water.
|
|
Figure 4.
Fig. 4. PEP-binding site. A, polar contacts of PEP inside
the active site. The C- trace and
side chains of the Cd^2+ enzyme are shown with salmon and white
bonds, respectively. Cd^2+ coordination is shown as transparent
bonds. PEP is shown as ball-and-sticks with gold bonds; the si
side of PEP is pointing up and to the left, and the re side is
pointing down and to the right. Blue, nitrogen; red, oxygen;
yellow, sulfur; pale blue, phosphorus; orange, carbon. B,
details of PEP geometry inside the active site. PEP is shown
with its si side pointing up. The angle between the plane
defined by C-2, C-1, and C-3 (blue triangle) and the plane
defined by C-1, C-3, and O-2 (yellow triangle) is ~12°,
indicating that the geometry of C-2 is intermediate between
trigonal and tetrahedral. WAT, water.
|
|
|
|
|
|
| |
The above figures are
reprinted
by permission from the ASBMB:
J Biol Chem
(2001,
276,
8393-8402)
copyright 2001.
|
|
| |
Figures were
selected
by an automated process.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
 |
 |
|
Literature references that cite this PDB file's key reference
|
|
|
| |
PubMed id
|
|
Reference
|
|
|
|
|
|
A.Roberts,
C.Furdui,
and
K.S.Anderson
(2010).
Observation of a chemically labile, noncovalent enzyme intermediate in the reaction of metal-dependent Aquifex pyrophilus KDO8PS by time-resolved mass spectrometry.
|
| |
Rapid Commun Mass Spectrom,
24,
1919-1924.
|
|
|
|
|
|
|
L.Cipolla,
L.Gabrielli,
D.Bini,
L.Russo,
and
N.Shaikh
(2010).
Kdo: a critical monosaccharide for bacteria viability.
|
| |
Nat Prod Rep,
27,
1618-1629.
|
|
|
|
|
|
|
U.Mamat,
H.Schmidt,
E.Munoz,
B.Lindner,
K.Fukase,
A.Hanuszkiewicz,
J.Wu,
T.C.Meredith,
R.W.Woodard,
R.Hilgenfeld,
J.R.Mesters,
and
O.Holst
(2009).
WaaA of the hyperthermophilic bacterium Aquifex aeolicus is a monofunctional 3-deoxy-D-manno-oct-2-ulosonic acid transferase involved in lipopolysaccharide biosynthesis.
|
| |
J Biol Chem,
284,
22248-22262.
|
|
|
|
|
|
|
M.Ahn,
A.L.Pietersma,
L.R.Schofield,
and
E.J.Parker
(2005).
Mechanistic divergence of two closely related aldol-like enzyme-catalysed reactions.
|
| |
Org Biomol Chem,
3,
4046-4049.
|
|
|
|
|
|
|
C.Sansom
(2001).
LPS inhibitors: key to overcoming multidrug-resistant bacteria?
|
| |
Drug Discov Today,
6,
499-500.
|
|
|
|
|
|
The most recent references are shown first.
Citation data come partly from CiteXplore and partly
from an automated harvesting procedure. Note that this is likely to be
only a partial list as not all journals are covered by
either method. However, we are continually building up the citation data
so more and more references will be included with time.
|
|



 Links
Links





