 |
PDBsum entry 5a1t

|
|
|
|
|
|

|

|

|

|
|

|
|

|
|

|
|

|
|

|
|

|
|
|
 |

|

|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Oxidoreductase
|
PDB id
|
|
|
|
5a1t
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|

|

|

|
|
|
|
|
|
|
|
|
Enzyme class:
|
|
E.C.1.1.1.27
- L-lactate dehydrogenase.
|
|
|
|
|
|
|
Reaction:
|
|
(S)-lactate + NAD+ = pyruvate + NADH + H+
|
|
|
|
|
|
(S)-lactate
Bound ligand (Het Group name = )
matches with 71.43% similarity
|
+
|
NAD(+)
Bound ligand (Het Group name = )
corresponds exactly
|
=
|
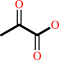 pyruvate
pyruvate
|
+
|
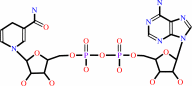 NADH
NADH
|
+
|
H(+)
|
|
|
|
|
|
|
|
|
|
|
|
|
Molecule diagrams generated from .mol files obtained from the
KEGG ftp site
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
| |
|
|
| |
|
DOI no:
|
Protein Sci
25:1319-1331
(2016)
|
|
PubMed id:
|
|
|
|
|
|
| |
|
Gradual neofunctionalization in the convergent evolution of trichomonad lactate and malate dehydrogenases.
|
|
P.A.Steindel,
E.H.Chen,
J.D.Wirth,
D.L.Theobald.
|
|
|
|
|
| |
ABSTRACT
|
|
|
|
| |
|
|
Lactate and malate dehydrogenases (LDH and MDH) are homologous, core metabolic
enzymes common to nearly all living organisms. LDHs have evolved convergently
from MDHs at least four times, achieving altered substrate specificity by a
different mechanism each time. For instance, the LDH of anaerobic trichomonad
parasites recently evolved independently from an ancestral trichomonad MDH by
gene duplication. LDH plays a central role in trichomonad metabolism by
catalyzing the reduction of pyruvate to lactate, thereby regenerating the NAD+
required for glycolysis. Using ancestral reconstruction methods, we identified
the biochemical and evolutionary mechanisms responsible for this convergent
event. The last common ancestor of these enzymes was a highly specific MDH,
similar to modern trichomonad MDHs. In contrast, the LDH lineage evolved
promiscuous activity by relaxing specificity in a gradual process of
neofunctionalization involving one highly detrimental substitution at the
"specificity residue" (R91L) and many additional mutations of small
effect. L91 has different functional consequences in LDHs and in MDHs,
indicating a prominent role for epistasis. Crystal structures of modern-day and
ancestral enzymes show that the evolution of substrate specificity paralleled
structural changes in dimerization and α-helix orientation. The relatively
small "specificity residue" of the trichomonad LDHs can accommodate a
range of substrate sizes and may permit solvent to access the active site, both
of which promote substrate promiscuity. The trichomonad LDHs present a
multi-faceted counterpoint to the independent evolution of LDHs in other
organisms and illustrate the diverse mechanisms by which protein function,
structure, and stability coevolve.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
 |
 |
|


 Links
Links











