 |
PDBsum entry 4zfa

|
|
|
|
|
|

|

|

|

|
|

|
|

|
|

|
|

|
|

|
|
|
 |

|

|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Oxidoreductase
|
PDB id
|
|
|
|
4zfa
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|

|

|

|
|
|
|
|
|
|
|
|
Enzyme class 2:
|
|
E.C.1.14.14.1
- unspecific monooxygenase.
|
|
|
|
|
|
|
Reaction:
|
|
an organic molecule + reduced [NADPH--hemoprotein reductase] + O2 = an alcohol + oxidized [NADPH--hemoprotein reductase] + H2O + H+
|
|
|
|
|
|
organic molecule
|
+
|
reduced [NADPH--hemoprotein reductase]
|
+
|
 O2
O2
|
=
|
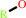 alcohol
alcohol
|
+
|
oxidized [NADPH--hemoprotein reductase]
|
+
|
H2O
|
+
|
H(+)
|
|
|
|
|
|
|
|
|
|
Cofactor:
|
|
Heme-thiolate
|
|
|
|
|
|
Enzyme class 3:
|
|
E.C.1.6.2.4
- NADPH--hemoprotein reductase.
|
|
|
|
|
|
|
Reaction:
|
|
2 oxidized [cytochrome P450] + NADPH = 2 reduced [cytochrome P450] + NADP+ + H+
|
|
|
|
|
|
2
×
oxidized [cytochrome P450]
|
+
|
 NADPH
NADPH
|
=
|
2
×
reduced [cytochrome P450]
|
+
|
 NADP(+)
NADP(+)
|
+
|
H(+)
|
|
|
|
|
|
|
|
|
|
Cofactor:
|
|
FAD; FMN
|
|
|
|
|
|
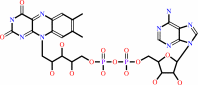 FAD
FAD
|
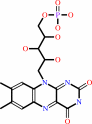 FMN
FMN
|
|
|
|
|
|
|
Note, where more than one E.C. class is given (as above), each may
correspond to a different protein domain or, in the case of polyprotein
precursors, to a different mature protein.
|
|
|
|
Molecule diagrams generated from .mol files obtained from the
KEGG ftp site
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
| |
|
|
| |
|
DOI no:
|
Biochemistry
55:3594-3606
(2016)
|
|
PubMed id:
|
|
|
|
|
|
| |
|
Effect of Mutation and Substrate Binding on the Stability of Cytochrome P450BM3 Variants.
|
|
I.Geronimo,
C.A.Denning,
W.E.Rogers,
T.Othman,
T.Huxford,
D.K.Heidary,
E.C.Glazer,
C.M.Payne.
|
|
|
|
|
| |
ABSTRACT
|
|
|
|
| |
|
|
Cytochrome P450BM3 is a heme-containing enzyme from Bacillus megaterium that
exhibits high monooxygenase activity and has a self-sufficient electron transfer
system in the full-length enzyme. Its potential synthetic applications drive
protein engineering efforts to produce variants capable of oxidizing nonnative
substrates such as pharmaceuticals and aromatic pollutants. However, promiscuous
P450BM3 mutants often exhibit lower stability, thereby hindering their
industrial application. This study demonstrated that the heme domain
R47L/F87V/L188Q/E267V/F81I pentuple mutant (PM) is destabilized because of the
disruption of hydrophobic contacts and salt bridge interactions. This was
directly observed from crystal structures of PM in the presence and absence of
ligands (palmitic acid and metyrapone). The instability of the tertiary
structure and heme environment of substrate-free PM was confirmed by pulse
proteolysis and circular dichroism, respectively. Binding of the inhibitor,
metyrapone, significantly stabilized PM, but the presence of the native
substrate, palmitic acid, had no effect. On the basis of high-temperature
molecular dynamics simulations, the lid domain, β-sheet 1, and Cys ligand loop
(a β-bulge segment connected to the heme) are the most labile regions and,
thus, potential sites for stabilizing mutations. Possible approaches to
stabilization include improvement of hydrophobic packing interactions in the lid
domain and introduction of new salt bridges into β-sheet 1 and the heme region.
An understanding of the molecular factors behind the loss of stability of
P450BM3 variants therefore expedites site-directed mutagenesis studies aimed at
developing thermostability.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
 |
 |
|



 Links
Links














