 |
PDBsum entry 4l9p
|
|
 |
|
|
|
|
|
|
Enzyme class 1:
|
|
Chains A, B:
E.C.2.5.1.58
- protein farnesyltransferase.
|
|
|
|
|
|
|
Reaction:
|
|
L-cysteinyl-[protein] + (2E,6E)-farnesyl diphosphate = S-(2E,6E)- farnesyl-L-cysteinyl-[protein] + diphosphate
|
|
|
|
|
|
L-cysteinyl-[protein]
Bound ligand (Het Group name = )
matches with 50.00% similarity
|
+
|
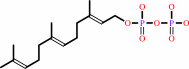 (2E,6E)-farnesyl diphosphate
(2E,6E)-farnesyl diphosphate
|
=
|
S-(2E,6E)- farnesyl-L-cysteinyl-[protein]
|
+
|
 diphosphate
diphosphate
|
|
|
|
|
|
|
|
|
|
Cofactor:
|
|
Mg(2+); Zn(2+)
|
|
|
|
|
|
Enzyme class 2:
|
|
Chain A:
E.C.2.5.1.59
- protein geranylgeranyltransferase type I.
|
|
|
|
|
|
|
Reaction:
|
|
geranylgeranyl diphosphate + L-cysteinyl-[protein] = S-geranylgeranyl-L- cysteinyl-[protein] + diphosphate
|
|
|
|
|
|
geranylgeranyl diphosphate
Bound ligand (Het Group name = )
matches with 43.24% similarity
|
+
|
L-cysteinyl-[protein]
|
=
|
S-geranylgeranyl-L- cysteinyl-[protein]
|
+
|
diphosphate
|
|
|
|
|
|
|
|
|
|
Cofactor:
|
|
Zn(2+)
|
|
|
|
|
|
|
|
|
Note, where more than one E.C. class is given (as above), each may
correspond to a different protein domain or, in the case of polyprotein
precursors, to a different mature protein.
|
|
|
|
Molecule diagrams generated from .mol files obtained from the
KEGG ftp site
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
| |
|
|
| |
|
DOI no:
|
Protein Sci
23:289-301
(2014)
|
|
PubMed id:
|
|
|
|
|
|
| |
|
Crystal structures of the fungal pathogen Aspergillus fumigatus protein farnesyltransferase complexed with substrates and inhibitors reveal features for antifungal drug design.
|
|
M.F.Mabanglo,
M.A.Hast,
N.B.Lubock,
H.W.Hellinga,
L.S.Beese.
|
|
|
|
|
| |
ABSTRACT
|
|
|
|
| |
|
|
Species of the fungal genus Aspergillus are significant human and agricultural
pathogens that are often refractory to existing antifungal treatments. Protein
farnesyltransferase (FTase), a critical enzyme in eukaryotes, is an attractive
potential target for antifungal drug discovery. We report high-resolution
structures of A. fumigatus FTase (AfFTase) in complex with substrates and
inhibitors. Comparison of structures with farnesyldiphosphate (FPP) bound in the
absence or presence of peptide substrate, corresponding to successive steps in
ordered substrate binding, revealed that the second substrate-binding step is
accompanied by motions of a loop in the catalytic site. Re-examination of other
FTase structures showed that this motion is conserved. The substrate- and
product-binding clefts in the AfFTase active site are wider than in human FTase
(hFTase). Widening is a consequence of small shifts in the α-helices that
comprise the majority of the FTase structure, which in turn arise from sequence
variation in the hydrophobic core of the protein. These structural effects are
key features that distinguish fungal FTases from hFTase. Their variation results
in differences in steady-state enzyme kinetics and inhibitor interactions and
presents opportunities for developing selective anti-fungal drugs by exploiting
size differences in the active sites. We illustrate the latter by comparing the
interaction of ED5 and Tipifarnib with hFTase and AfFTase. In AfFTase, the wider
groove enables ED5 to bind in the presence of FPP, whereas in hFTase it binds
only in the absence of substrate. Tipifarnib binds similarly to both enzymes but
makes less extensive contacts in AfFTase with consequently weaker binding.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
 |
 |
|
|




 Links
Links














