 |
PDBsum entry 4fns
|
|

|
|
PDB id:
|
|
|
|
| Name: |
|
Hydrolase
|
|
|
Title:
|
|
Crystal structure of gh36 alpha-galactosidase agaa a355e from geobacillus stearothermophilus in complex with 1- deoxygalactonojirimycin
|
|
Structure:
|
|
Alpha-galactosidase agaa. Chain: a, b, c, d. Engineered: yes. Mutation: yes
|
|
Source:
|
|
Geobacillus stearothermophilus. Organism_taxid: 1422. Gene: agaa. Expressed in: escherichia coli. Expression_system_taxid: 562
|
|
Resolution:
|
|
|
2.60Å
|
R-factor:
|
0.177
|
R-free:
|
0.231
|
|
|
Authors:
|
|
R.Merceron,M.Foucault,R.Haser,R.Mattes,H.Watzlawick,P.Gouet
|
|
Key ref:
|
|
R.Merceron
et al.
(2012).
The molecular mechanism of thermostable α-galactosidases AgaA and AgaB explained by x-ray crystallography and mutational studies.
J Biol Chem,
287,
39642-39652.
PubMed id:
|
|
|
Date:
|
|
|
20-Jun-12
|
Release date:
|
03-Oct-12
|
|
|
|
|
|
|
PROCHECK
|

|
|
|
|
|
Headers
|
|
|
|
References
|
|
|
|
|
|

|
|
Q9ALJ4
(AGAA_GEOSE) -
Alpha-galactosidase AgaA from Geobacillus stearothermophilus
|
|
|
|
Seq:
Struc:
|
|
|
|
729 a.a.
718 a.a.*
|
|

|

|
|
|
|
|
|
|
|
|
|
|
|
Key: |
 |
PfamA domain |
|
|
 |
Secondary structure |
|
 |
CATH domain |
|
|
*
PDB and UniProt seqs differ
at 3 residue positions (black
crosses)
|
|
|
|
|
|
|
|
|
|
|
|
|
Enzyme class:
|
|
E.C.3.2.1.22
- alpha-galactosidase.
|
|
|
|
|
|
|
Reaction:
|
|
Melibiose + H2O = galactose + glucose
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Cofactor:
|
|
Mg(2+); NAD(+)
|
|
|
|
|
|
Mg(2+)
|
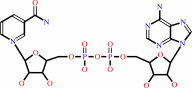 NAD(+)
NAD(+)
|
|
|
|
|
|
|
Molecule diagrams generated from .mol files obtained from the
KEGG ftp site
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
| |
|
|
| |
|
|
J Biol Chem
287:39642-39652
(2012)
|
|
PubMed id:
|
|
|
|
|
|
| |
|
The molecular mechanism of thermostable α-galactosidases AgaA and AgaB explained by x-ray crystallography and mutational studies.
|
|
R.Merceron,
M.Foucault,
R.Haser,
R.Mattes,
H.Watzlawick,
P.Gouet.
|
|
|
|
|
| |
ABSTRACT
|
|
|
|
| |
|
|
The α-galactosidase AgaA from the thermophilic microorganism Geobacillus
stearothermophilus has great industrial potential because it is fully active at
338 K against raffinose and can increase the yield of manufactured sucrose. AgaB
has lower affinity for its natural substrates but is a powerful tool for the
enzymatic synthesis of disaccharides by transglycosylation. These two enzymes
have 97% identity and belong to the glycoside hydrolase (GH) family GH36 for
which few structures are available. To understand the structural basis
underlying the differences between these two enzymes, we determined the crystal
structures of AgaA and AgaB by molecular replacement at 3.2- and 1.8
Å-resolution, respectively. We also solved a 2.8-Å structure of the
AgaA(A355E) mutant, which has enzymatic properties similar to those of AgaB. We
observe that residue 355 is located 20 Å away from the active site and that
the A355E substitution causes structural rearrangements resulting in a
significant displacement of the invariant Trp(336) at catalytic subsite -1.
Hence, the active cleft of AgaA is narrowed in comparison with AgaB, and AgaA is
more efficient than AgaB against its natural substrates. The structure of
AgaA(A355E) complexed with 1-deoxygalactonojirimycin reveals an induced fit
movement; there is a rupture of the electrostatic interaction between Glu(355)
and Asn(335) and a return of Trp(336) to an optimal position for ligand
stacking. The structures of two catalytic mutants of AgaA(A355E) complexed with
raffinose and stachyose show that the binding interactions are stronger at
subsite -1 to enable the binding of various α-galactosides.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
 |
 |
|


 Links
Links






