 |
PDBsum entry 4blc

|
|
|
|
|
|

|

|

|

|
|

|
|

|
|

|
|

|
|

|
|
|
 |

|

|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Oxidoreductase
|
PDB id
|
|
|
|
4blc
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|

|

|

|
|
|
|
|
|
|
|
 |
Contents |
 |
|
|
|
|
|
|
|
|
|
|
|
|
* Residue conservation analysis
|
|
|
|
|
|
PDB id:
|
|
|
|
| Name: |
|
Oxidoreductase
|
|
|
Title:
|
|
The structure of orthorhombic crystals of beef liver catalase
|
|
Structure:
|
|
Protein (catalase). Chain: a, b, c, d. Other_details: each subunit contains a heme and an NADP
|
|
Source:
|
|
Bos taurus. Cattle. Organism_taxid: 9913. Organ: liver. Other_details: purchased from sigma chemical co., St.Louis, mo
|
|
Biol. unit:
|
|
Tetramer (from
)
|
|
Resolution:
|
|
|
2.30Å
|
R-factor:
|
0.205
|
R-free:
|
0.273
|
|
|
Authors:
|
|
T.P.Ko,J.Day,A.Malkin,A.Mcpherson
|
Key ref:

|
|
T.P.Ko
et al.
(1999).
Structure of orthorhombic crystals of beef liver catalase.
Acta Crystallogr D Biol Crystallogr,
55,
1383-1394.
PubMed id:
DOI:

|
|
|
Date:
|
|
|
27-Sep-98
|
Release date:
|
14-Oct-98
|
|
|
|
|
|
|
PROCHECK
|

|
|
|
|
|
Headers
|
|
|
|
References
|
|
|
|
|
|

|
|
P00432
(CATA_BOVIN) -
Catalase from Bos taurus
|
|
|
|
Seq:
Struc:
|
|
|
|
527 a.a.
499 a.a.*
|
|

|

|
|
|
|
|
|
|
|
|
|
|
|
Key: |
 |
PfamA domain |
|
|
 |
Secondary structure |
|
 |
CATH domain |
|
|
*
PDB and UniProt seqs differ
at 2 residue positions (black
crosses)
|
|
|
|
|
|
|
|
|
|
|
|
|
Enzyme class:
|
|
E.C.1.11.1.6
- catalase.
|
|
|
|
|
|
|
Reaction:
|
|
2 H2O2 = O2 + 2 H2O
|
|
|
|
|
|
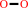 2
×
H2O2
2
×
H2O2
|
=
|
 O2
O2
|
+
|
2
×
H2O
|
|
|
|
|
|
|
|
|
|
Cofactor:
|
|
Heme; Mn(2+)
|
|
|
|
|
|
Heme
Bound ligand (Het Group name =
HEM)
matches with 95.45% similarity
|
Mn(2+)
|
|
|
|
|
|
|
Molecule diagrams generated from .mol files obtained from the
KEGG ftp site
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
| |
|
|
| |
|
DOI no:
|
Acta Crystallogr D Biol Crystallogr
55:1383-1394
(1999)
|
|
PubMed id:
|
|
|
|
|
|
| |
|
Structure of orthorhombic crystals of beef liver catalase.
|
|
T.P.Ko,
J.Day,
A.J.Malkin,
A.McPherson.
|
|
|
|
|
| |
ABSTRACT
|
|
|
|
| |
|
|
The growth mechanisms and physical properties of the orthorhombic crystal form
of beef liver catalase were investigated using in situ atomic force microscopy
(AFM). It was observed that the crystals grow in the <001> direction by an
unusual progression of sequential two-dimensional nuclei of half unit-cell
layers corresponding to the 'bottoms' and 'tops' of unit cells. These were
easily discriminated by their alternating asymmetric shapes and their strong
growth-rate anisotropy. This pattern has not previously been observed with other
macromolecular crystals. Orthorhombic beef liver catalase crystals exhibit an
extremely high defect density and incorporate great numbers of misoriented
microcrystals, revealed intact by etching experiments, which may explain their
marginal diffraction properties. To facilitate interpretation of AFM results in
terms of intermolecular interactions, the structure of the orthorhombic
crystals, having an entire tetramer of the enzyme as the asymmetric unit, was
solved by molecular replacement using a model derived from a trigonal crystal
form. It was subsequently refined by conventional techniques. Although the
packing of molecules in the two unit cells was substantially different, with
very few exceptions no significant differences in the molecular structures were
observed. In addition, no statistically significant deviation from ideal 222
molecular symmetry appeared within the tetramer. The packing of molecules in the
crystal revealed by X-ray analysis explained in a satisfying way the process of
crystal growth revealed by AFM.
|
|
|
|
|
|
| |
Selected figure(s)
|
|
|
|
| |
|
|

|
|

|
|
Figure 4.
Figure 4 A 5 × 5 µm atomic force micrograph of the
(001) surface of an orthorhombic beef liver catalase crystal
showing step edges and two-dimensional islands. The roughness of
the edges suggests the presence and incorporation of high levels
of macromolecular impurities which retard normal step
advancement at sites of addition.
|
|
Figure 9.
Figure 9 A 42 × 42 µm area recorded by AFM on the
(001) surface of an orthorhombic catalase crystal which has been
slightly etched near equilibrium conditions. This procedure
reveals defects and incorporated impurities including
incorporated microcrystals which are misoriented with respect to
the underlying lattice. Here, it can be seen that there is a
striking number of such misoriented microcrystals dispersed
throughout the crystal, some as long as 10 µm.
|
|
|
|
|
|
| |
The above figures are
reprinted
by permission from the IUCr:
Acta Crystallogr D Biol Crystallogr
(1999,
55,
1383-1394)
copyright 1999.
|
|
| |
Figures were
selected
by an automated process.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
 |
 |
|
Literature references that cite this PDB file's key reference
|
|
|
| |
PubMed id
|
|
Reference
|
|
|
|
|
|
D.A.Svistunenko,
and
G.A.Jones
(2009).
Tyrosyl radicals in proteins: a comparison of empirical and density functional calculated EPR parameters.
|
| |
Phys Chem Chem Phys,
11,
6600-6613.
|
|
|
|
|
|
|
D.A.Svistunenko,
and
C.E.Cooper
(2004).
A new method of identifying the site of tyrosyl radicals in proteins.
|
| |
Biophys J,
87,
582-595.
|
|
|
|
|
|
|
J.Wang,
M.Ortiz-Maldonado,
B.Entsch,
V.Massey,
D.Ballou,
and
D.L.Gatti
(2002).
Protein and ligand dynamics in 4-hydroxybenzoate hydroxylase.
|
| |
Proc Natl Acad Sci U S A,
99,
608-613.
|
|
|
PDB codes:
|
|
|
|
|
|
|
The most recent references are shown first.
Citation data come partly from CiteXplore and partly
from an automated harvesting procedure. Note that this is likely to be
only a partial list as not all journals are covered by
either method. However, we are continually building up the citation data
so more and more references will be included with time.
Where a reference describes a PDB structure, the PDB
codes are
shown on the right.
|
|


 Links
Links






