 |
PDBsum entry 3f4b

|
|
|
|
|
|

|

|

|

|
|

|
|

|
|

|
|

|
|

|
|

|
|
|
 |

|

|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Oxidoreductase
|
PDB id
|
|
|
|
3f4b
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|

|

|

|
|
|
|
|
|
|
|
 |
Contents |
 |
|
|
|
|
|
|
|
|
|
|
|
|
* Residue conservation analysis
|
|
|
|
|
|
|
|
|
|
|
Enzyme class:
|
|
E.C.1.3.1.9
- enoyl-[acyl-carrier-protein] reductase (NADH).
|
|
|
|
|
|
|
Reaction:
|
|
a 2,3-saturated acyl-[ACP] + NAD+ = a (2E)-enoyl-[ACP] + NADH + H+
|
|
|
|
|
|
2,3-saturated acyl-[ACP]
|
+
|
NAD(+)
Bound ligand (Het Group name = )
corresponds exactly
|
=
|
(2E)-enoyl-[ACP]
|
+
|
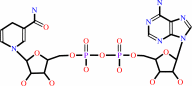 NADH
NADH
|
+
|
H(+)
|
|
|
|
|
|
|
|
|
|
|
|
|
Molecule diagrams generated from .mol files obtained from the
KEGG ftp site
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
| |
|
|
| |
|
|
Cell Host Microbe
4:567-578
(2008)
|
|
PubMed id:
|
|
|
|
|
|
| |
|
The fatty acid biosynthesis enzyme FabI plays a key role in the development of liver-stage malarial parasites.
|
|
M.Yu,
T.R.Kumar,
L.J.Nkrumah,
A.Coppi,
S.Retzlaff,
C.D.Li,
B.J.Kelly,
P.A.Moura,
V.Lakshmanan,
J.S.Freundlich,
J.C.Valderramos,
C.Vilcheze,
M.Siedner,
J.H.Tsai,
B.Falkard,
A.B.Sidhu,
L.A.Purcell,
P.Gratraud,
L.Kremer,
A.P.Waters,
G.Schiehser,
D.P.Jacobus,
C.J.Janse,
A.Ager,
W.R.Jacobs,
J.C.Sacchettini,
V.Heussler,
P.Sinnis,
D.A.Fidock.
|
|
|
|
|
| |
ABSTRACT
|
|
|
|
| |
|
|
The fatty acid synthesis type II pathway has received considerable interest as a
candidate therapeutic target in Plasmodium falciparum asexual blood-stage
infections. This apicoplast-resident pathway, distinct from the mammalian type I
process, includes FabI. Here, we report synthetic chemistry and transfection
studies concluding that Plasmodium FabI is not the target of the antimalarial
activity of triclosan, an inhibitor of bacterial FabI. Disruption of fabI in P.
falciparum or the rodent parasite P. berghei does not impede blood-stage growth.
In contrast, mosquito-derived, FabI-deficient P. berghei sporozoites are
markedly less infective for mice and typically fail to complete liver-stage
development in vitro. This defect is characterized by an inability to form
intrahepatic merosomes that normally initiate blood-stage infections. These data
illuminate key differences between liver- and blood-stage parasites in their
requirements for host versus de novo synthesized fatty acids, and create new
prospects for stage-specific antimalarial interventions.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
 |
 |
|
Literature references that cite this PDB file's key reference
|
|
|
| |
PubMed id
|
|
Reference
|
|
|
|
|
|
G.Plata,
T.L.Hsiao,
K.L.Olszewski,
M.Llinás,
and
D.Vitkup
(2010).
Reconstruction and flux-balance analysis of the Plasmodium falciparum metabolic network.
|
| |
Mol Syst Biol,
6,
408.
|
|
|
|
|
|
|
K.L.Olszewski,
M.W.Mather,
J.M.Morrisey,
B.A.Garcia,
A.B.Vaidya,
J.D.Rabinowitz,
and
M.Llinás
(2010).
Branched tricarboxylic acid metabolism in Plasmodium falciparum.
|
| |
Nature,
466,
774-778.
|
|
|
|
|
|
|
D.Mazier,
L.Rénia,
and
G.Snounou
(2009).
A pre-emptive strike against malaria's stealthy hepatic forms.
|
| |
Nat Rev Drug Discov,
8,
854-864.
|
|
|
|
|
|
The most recent references are shown first.
Citation data come partly from CiteXplore and partly
from an automated harvesting procedure. Note that this is likely to be
only a partial list as not all journals are covered by
either method. However, we are continually building up the citation data
so more and more references will be included with time.
|
|


 Links
Links











