 |
PDBsum entry 3cmc

|
|
|
|
|
|

|

|

|

|
|

|
|

|
|

|
|

|
|

|
|
|
 |

|

|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Oxidoreductase
|
PDB id
|
|
|
|
3cmc
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|

|

|

|
|
|
|
|
|
|
|
 |
Contents |
 |
|
|
|
|
|
|
|
|
|
|
|
|
* Residue conservation analysis
|
|
|
|
|
|
|
|
|
|
|
Enzyme class:
|
|
E.C.1.2.1.12
- glyceraldehyde-3-phosphate dehydrogenase (phosphorylating).
|
|
|
|
|
|
|
Pathway:
|
|
Glyceraldehyde-3-phosphate Dehydrogenase (phosphorylating)
|
|
|
|
|
|
Reaction:
|
|
D-glyceraldehyde 3-phosphate + phosphate + NAD+ = (2R)-3-phospho- glyceroyl phosphate + NADH + H+
|
|
|
|
|
|
D-glyceraldehyde 3-phosphate
Bound ligand (Het Group name = )
corresponds exactly
|
+
|
 phosphate
phosphate
|
+
|
NAD(+)
Bound ligand (Het Group name = )
corresponds exactly
|
=
|
(2R)-3-phospho- glyceroyl phosphate
|
+
|
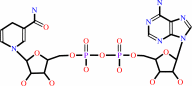 NADH
NADH
|
+
|
H(+)
|
|
|
|
|
|
|
|
|
|
|
|
|
Molecule diagrams generated from .mol files obtained from the
KEGG ftp site
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
| |
|
|
| |
|
DOI no:
|
J Biol Chem
283:21693-21702
(2008)
|
|
PubMed id:
|
|
|
|
|
|
| |
|
Trapping of the Thioacylglyceraldehyde-3-phosphate Dehydrogenase Intermediate from Bacillus stearothermophilus: DIRECT EVIDENCE FOR A FLIP-FLOP MECHANISM.
|
|
S.Moniot,
S.Bruno,
C.Vonrhein,
C.Didierjean,
S.Boschi-Muller,
M.Vas,
G.Bricogne,
G.Branlant,
A.Mozzarelli,
C.Corbier.
|
|
|
|
|
| |
ABSTRACT
|
|
|
|
| |
|
|
The crystal structure of the thioacylenzyme intermediate of the phosphorylating
glyceraldehyde-3-phosphate dehydrogenase (GAPDH) from Bacillus
stearothermophilus has been solved at 1.8A resolution. Formation of the
intermediate was obtained by diffusion of the natural substrate within the
crystal of the holoenzyme in the absence of inorganic phosphate. To define the
soaking conditions suitable for the isolation and accumulation of the
intermediate, a microspectrophotometric characterization of the reaction of
GAPDH in single crystals was carried out, following NADH formation at 340 nm.
When compared with the structure of the Michaelis complex ( Didierjean, C.,
Corbier, C., Fatih, M., Favier, F., Boschi-Muller, S., Branlant, G., and Aubry,
A. (2003) J. Biol. Chem. 278, 12968-12976 ) the 206-210 loop is shifted and now
forms part of the so-called "new P(i)" site. The locations of both the O1 atom
and the C3-phosphate group of the substrate are also changed. Altogether, the
results provide evidence for the flipping of the C3-phosphate group occurring
concomitantly or after the redox step.
|
|
|
|
|
|
| |
Selected figure(s)
|
|
|
|
| |
|
|

|
|

|
|
Figure 4.
FIGURE 4. Alternate conformations of the 206-212 loop and
re-location of the P[i] anion binding site. A, stereoscopic view
of the O subunit region spanning residues 205-218. The 206-212
loop is represented in two alternate conformations: in dark
gray, the classic conformation delineates the P[i] anion binding
site where a sulfate is bound (conformation A). In light gray,
the alternate conformation exhibits a shift toward the C3P of
the intermediate (conformation B). This motion allows Thr-208
and Gly-209 to participate to the formation of the new P[i] site
and thus to the binding of the C3P group of the intermediate. B,
superimposition of the 206-212 loop with representative GAPDH
structures. The superimposition of the 206-212 loop under its
classic conformation (dark gray) with the holo-structure from B.
stearothermophilus (in white, pdb code 1gd1 (10)) is reported on
the left, and the superimposition of the 206-212 loop under its
alternate conformation with the structure of T. maritima GAPDH
(in white, pdb code 1hdg (11)) is reported on the right. All
structures are represented in schematic mode with their
corresponding bound anion in stick mode. To facilitate
comparison, both conformations of our model were represented in
each superimposition, shading the one that is not considered.
|
|
Figure 5.
FIGURE 5. Structural scenario proposed for the catalytic
mechanism. Only hydrogen atoms directly involved in the
catalytic mechanism are represented. G3P initially binds to the
active site of GAPDH with its C3P group located in the P[s]
site. Reaction begins through the nucleophilic attack of the
Cys-149 thiolate function on the aldehydic carbon C1 of the
substrate (step 1), which leads to a tetrahedral intermediate
called hemithioacetal (HTA). This step is followed by hydride
transfer (step 2), assisted by His-176, from the C1 atom of G3P
to the C4 atom of NAD^+. At this stage, C3P is thought to be
located in the P[s] site, and the O1 atom of G3P is expected to
interact with His-176 (N  ) (see "Discussion"). To
allow NADH release, the substrate must undergo a conformational
change during which the orientation of the O1 atom changes while
the C3P group flips toward the new P[i] site (step 3). It
results in the loss of the interaction between the substrate and
the cofactor required for NADH release (TAE: thioacylenzyme
intermediate corresponding to the structure depicted here).
NAD^+ enters the active site and likely promotes the relocation
of O1 and C3P to their initial position. The P[i] site (either
in the classic or new position, indicated as "Pi?") is free to
bind an inorganic phosphate for the nucleophilic attack on the
thioacylenzyme. This last step is also assisted by His-176 and
leads to the formation of 1,3-bisphosphoglycerate. Subsequent
release of the product leaves the enzyme in its holo-state,
ready to accommodate a new G3P molecule. ) (see "Discussion"). To
allow NADH release, the substrate must undergo a conformational
change during which the orientation of the O1 atom changes while
the C3P group flips toward the new P[i] site (step 3). It
results in the loss of the interaction between the substrate and
the cofactor required for NADH release (TAE: thioacylenzyme
intermediate corresponding to the structure depicted here).
NAD^+ enters the active site and likely promotes the relocation
of O1 and C3P to their initial position. The P[i] site (either
in the classic or new position, indicated as "Pi?") is free to
bind an inorganic phosphate for the nucleophilic attack on the
thioacylenzyme. This last step is also assisted by His-176 and
leads to the formation of 1,3-bisphosphoglycerate. Subsequent
release of the product leaves the enzyme in its holo-state,
ready to accommodate a new G3P molecule.
|
|
|
|
|
|
| |
The above figures are
reprinted
by permission from the ASBMB:
J Biol Chem
(2008,
283,
21693-21702)
copyright 2008.
|
|
| |
Figures were
selected
by an automated process.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
 |
 |
|
Literature references that cite this PDB file's key reference
|
|
|
| |
PubMed id
|
|
Reference
|
|
|
|
|
|
O.S.Smart,
T.O.Womack,
C.Flensburg,
P.Keller,
W.Paciorek,
A.Sharff,
C.Vonrhein,
and
G.Bricogne
(2012).
Exploiting structure similarity in refinement: automated NCS and target-structure restraints in BUSTER.
|
| |
Acta Crystallogr D Biol Crystallogr,
68,
368-380.
|
|
|
PDB codes:
|
|
|
|
|
|
|
|
W.J.Cook,
O.Senkovich,
and
D.Chattopadhyay
(2009).
An unexpected phosphate binding site in glyceraldehyde 3-phosphate dehydrogenase: crystal structures of apo, holo and ternary complex of Cryptosporidium parvum enzyme.
|
| |
BMC Struct Biol,
9,
9.
|
|
|
PDB codes:
|
|
|
|
|
|
|
The most recent references are shown first.
Citation data come partly from CiteXplore and partly
from an automated harvesting procedure. Note that this is likely to be
only a partial list as not all journals are covered by
either method. However, we are continually building up the citation data
so more and more references will be included with time.
Where a reference describes a PDB structure, the PDB
codes are
shown on the right.
|
|


 Links
Links












