 |
PDBsum entry 3g4a

|
|
|
|
|
 |
Contents |
 |
|
|
|
|
|
|
|
|
|
|
|
|
* Residue conservation analysis
|
|
|
|
|
|
PDB id:
|
|
|
|
| Name: |
|
Transferase
|
|
|
Title:
|
|
Crystal structure of flavine dependant thymidylate synthase s88a mutant from thermotoga maritima at 1.95 angstrom resolution
|
|
Structure:
|
|
Thymidylate synthase thyx. Chain: a, b, c, d. Synonym: ts, tsase. Engineered: yes. Mutation: yes
|
|
Source:
|
|
Thermotoga maritima msb8. Organism_taxid: 243274. Strain: msb8 / dsm 3109 / jcm 10099. Atcc: 43589. Gene: thy1, thyx, tm_0449. Expressed in: escherichia coli. Expression_system_taxid: 562.
|
|
Resolution:
|
|
|
1.95Å
|
R-factor:
|
0.187
|
R-free:
|
0.222
|
|
|
Authors:
|
|
I.I.Mathews,S.A.Lesley,A.Kohen
|
Key ref:

|
|
E.M.Koehn
et al.
(2009).
An unusual mechanism of thymidylate biosynthesis in organisms containing the thyX gene.
Nature,
458,
919-923.
PubMed id:
DOI:

|
|
|
Date:
|
|
|
03-Feb-09
|
Release date:
|
07-Apr-09
|
|
|
|
|
|
|
PROCHECK
|

|
|
|
|
|
Headers
|
|
|
|
References
|
|
|
|
|
|

|
|
Q9WYT0
(THYX_THEMA) -
Flavin-dependent thymidylate synthase from Thermotoga maritima (strain ATCC 43589 / DSM 3109 / JCM 10099 / NBRC 100826 / MSB8)
|
|
|
|
Seq:
Struc:
|
|
|
|
220 a.a.
218 a.a.*
|
|

|

|
|
|
|
|
|
|
|
|
|
|
|
Key: |
 |
PfamA domain |
|
|
 |
Secondary structure |
|
 |
CATH domain |
|
|
*
PDB and UniProt seqs differ
at 1 residue position (black
cross)
|
|
|
|
|
|
|
|
|
|
|
|
|
Enzyme class:
|
|
E.C.2.1.1.148
- thymidylate synthase (FAD).
|
|
|
|
|
|
|
Reaction:
|
|
dUMP + (6R)-5,10-methylene-5,6,7,8-tetrahydrofolate + NADPH + H+ = dTMP + (6S)-5,6,7,8-tetrahydrofolate + NADP+
|
|
|
|
|
|
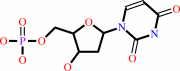 dUMP
dUMP
|
+
|
(6R)-5,10-methylene-5,6,7,8-tetrahydrofolate
Bound ligand (Het Group name = )
corresponds exactly
|
+
|
 NADPH
NADPH
|
+
|
H(+)
Bound ligand (Het Group name = )
matches with 71.19% similarity
|
=
|
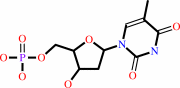 dTMP
dTMP
|
+
|
(6S)-5,6,7,8-tetrahydrofolate
|
+
|
 NADP(+)
NADP(+)
|
|
|
|
|
|
|
|
|
|
|
|
|
Molecule diagrams generated from .mol files obtained from the
KEGG ftp site
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
| |
|
|
| |
|
DOI no:
|
Nature
458:919-923
(2009)
|
|
PubMed id:
|
|
|
|
|
|
| |
|
An unusual mechanism of thymidylate biosynthesis in organisms containing the thyX gene.
|
|
E.M.Koehn,
T.Fleischmann,
J.A.Conrad,
B.A.Palfey,
S.A.Lesley,
I.I.Mathews,
A.Kohen.
|
|
|
|
|
| |
ABSTRACT
|
|
|
|
| |
|
|
Biosynthesis of the DNA base thymine depends on activity of the enzyme
thymidylate synthase to catalyse the methylation of the uracil moiety of
2'-deoxyuridine-5'-monophosphate. All known thymidylate synthases rely on an
active site residue of the enzyme to activate 2'-deoxyuridine-5'-monophosphate.
This functionality has been demonstrated for classical thymidylate synthases,
including human thymidylate synthase, and is instrumental in mechanism-based
inhibition of these enzymes. Here we report an example of thymidylate
biosynthesis that occurs without an enzymatic nucleophile. This unusual
biosynthetic pathway occurs in organisms containing the thyX gene, which codes
for a flavin-dependent thymidylate synthase (FDTS), and is present in several
human pathogens. Our findings indicate that the putative active site nucleophile
is not required for FDTS catalysis, and no alternative nucleophilic residues
capable of serving this function can be identified. Instead, our findings
suggest that a hydride equivalent (that is, a proton and two electrons) is
transferred from the reduced flavin cofactor directly to the uracil ring,
followed by an isomerization of the intermediate to form the product,
2'-deoxythymidine-5'-monophosphate. These observations indicate a very different
chemical cascade than that of classical thymidylate synthases or any other known
biological methylation. The findings and chemical mechanism proposed here,
together with available structural data, suggest that selective inhibition of
FDTSs, with little effect on human thymine biosynthesis, should be feasible.
Because several human pathogens depend on FDTS for DNA biosynthesis, its unique
mechanism makes it an attractive target for antibiotic drugs.
|
|
|
|
|
|
| |
Selected figure(s)
|
|
|
|
| |
|
|

|
|

|
|
Figure 1.
Figure 1: Thymidylate synthase mechanisms. a, The chemical
mechanism for the classical thymidylate synthase catalysed
reaction^1, ^2. b, The chemical mechanism for the FDTS proposed
hitherto^11. c, The newly proposed mechanism for the FDTS that
does not rely on an enzymatic nucleophile. The conserved
enzymatic nucleophile is orange, the methylene is purple, the
reducing hydride from H[4]folate is green, and the hydride from
FADH[2] is red. R = 2'-deoxyribose-5'-phosphate; R' =
(p-aminobenzoyl)-glutamate; R'' =
adenosine-5'-pyroposphate-ribityl.
|
|
Figure 2.
Figure 2: Crystal structures of the FDTS–FAD–dUMP complex.
a, Wild-type TmFDTS (Protein Data Bank 1o26); b, S88A mutant
(Protein Data Bank 3g4a); c, S88C mutant (Protein Data Bank
3g4c). The distance between the C6 carbon of dUMP and the
reducing centre of the flavin (N5 of FAD) is 3.4 Å for all
three enzymes. The distances of the side chain of residue 88 to
C6 are 4.3, 4.5 and 4.1 Å, for wild-type FDTS, S88A and
S88C, respectively. The electron density maps are 2F[o] - F[c]
with a contour level of 1.0 sigma.
|
|
|
|
|
|
| |
The above figures are
reprinted
from an Open Access publication published by Macmillan Publishers Ltd:
Nature
(2009,
458,
919-923)
copyright 2009.
|
|
| |
Figures were
selected
by an automated process.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
 |
 |
|
Literature references that cite this PDB file's key reference
|
|
|
| |
PubMed id
|
|
Reference
|
|
|
|
|
|
F.Yan,
and
D.G.Fujimori
(2011).
RNA methylation by Radical SAM enzymes RlmN and Cfr proceeds via methylene transfer and hydride shift.
|
| |
Proc Natl Acad Sci U S A,
108,
3930-3934.
|
|
|
|
|
|
|
A.E.Speers,
and
B.F.Cravatt
(2010).
Ligands in crystal structures that aid in functional characterization.
|
| |
Acta Crystallogr Sect F Struct Biol Cryst Commun,
66,
1306-1308.
|
|
|
|
|
|
|
D.Weekes,
S.S.Krishna,
C.Bakolitsa,
I.A.Wilson,
A.Godzik,
and
J.Wooley
(2010).
TOPSAN: a collaborative annotation environment for structural genomics.
|
| |
BMC Bioinformatics,
11,
426.
|
|
|
|
|
|
|
E.M.Koehn,
and
A.Kohen
(2010).
Flavin-dependent thymidylate synthase: a novel pathway towards thymine.
|
| |
Arch Biochem Biophys,
493,
96.
|
|
|
|
|
|
|
E.J.Loveridge,
G.Maglia,
and
R.K.Allemann
(2009).
The role of arginine 28 in catalysis by dihydrofolate reductase from the hyperthermophile Thermotoga maritima.
|
| |
Chembiochem,
10,
2624-2627.
|
|
|
|
|
|
|
M.P.Costi,
and
S.Ferrari
(2009).
Biochemistry: Anchors away.
|
| |
Nature,
458,
840-841.
|
|
|
|
|
|
|
Y.Zhang,
I.Thiele,
D.Weekes,
Z.Li,
L.Jaroszewski,
K.Ginalski,
A.M.Deacon,
J.Wooley,
S.A.Lesley,
I.A.Wilson,
B.Palsson,
A.Osterman,
and
A.Godzik
(2009).
Three-dimensional structural view of the central metabolic network of Thermotoga maritima.
|
| |
Science,
325,
1544-1549.
|
|
|
|
|
|
|
Z.Wang,
A.Chernyshev,
E.M.Koehn,
T.D.Manuel,
S.A.Lesley,
and
A.Kohen
(2009).
Oxidase activity of a flavin-dependent thymidylate synthase.
|
| |
FEBS J,
276,
2801-2810.
|
|
|
|
|
|
The most recent references are shown first.
Citation data come partly from CiteXplore and partly
from an automated harvesting procedure. Note that this is likely to be
only a partial list as not all journals are covered by
either method. However, we are continually building up the citation data
so more and more references will be included with time.
|
|


 Links
Links




