
|
|
|
|
|
 |
Contents |
 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
* Residue conservation analysis
|
|
|
|
|
|
|
|
|
|
Enzyme class:
|
|
Chain A:
E.C.1.18.1.1
- rubredoxin--NAD(+) reductase.
|
|
|
|
|
|
|
Reaction:
|
|
2 reduced [rubredoxin] + NAD+ + H+ = 2 oxidized [rubredoxin] + NADH
|
|
|
|
|
|
2
×
reduced [rubredoxin]
|
+
|
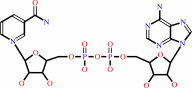 NAD(+)
NAD(+)
|
+
|
H(+)
|
=
|
2
×
oxidized [rubredoxin]
|
+
|
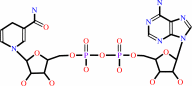 NADH
NADH
|
|
|
|
|
|
|
|
|
|
Cofactor:
|
|
FAD; Fe cation
|
|
|
|
|
|
FAD
Bound ligand (Het Group name =
FAD)
corresponds exactly
|
Fe cation
|
|
|
|
|
|
|
Molecule diagrams generated from .mol files obtained from the
KEGG ftp site
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
| |
|
|
| |
|
DOI no:
|
Proc Natl Acad Sci U S A
104:12276-12281
(2007)
|
|
PubMed id:
|
|
|
|
|
|
| |
|
Crystal structure of the electron transfer complex rubredoxin rubredoxin reductase of Pseudomonas aeruginosa.
|
|
G.Hagelueken,
L.Wiehlmann,
T.M.Adams,
H.Kolmar,
D.W.Heinz,
B.Tümmler,
W.D.Schubert.
|
|
|
|
|
| |
ABSTRACT
|
|
|
|
| |
|
|
Crude oil spills represent a major ecological threat because of the chemical
inertness of the constituent n-alkanes. The Gram-negative bacterium Pseudomonas
aeruginosa is one of the few bacterial species able to metabolize such
compounds. Three chromosomal genes, rubB, rubA1, and rubA2 coding for an
NAD(P)H:rubredoxin reductase (RdxR) and two rubredoxins (Rdxs) are indispensable
for this ability. They constitute an electron transport (ET) pathway that
shuttles reducing equivalents from carbon metabolism to the membrane-bound
alkane hydroxylases AlkB1 and AlkB2. The RdxR-Rdx system also is crucial as part
of the oxidative stress response in archaea or anaerobic bacteria. The redox
couple has been analyzed in detail as a model system for ET processes. We have
solved the structure of RdxR of P. aeruginosa both alone and in complex with
Rdx, without the need for cross-linking, and both structures were refined at
2.40- and 2.45-A resolution, respectively. RdxR consists of two cofactor-binding
domains and a C-terminal domain essential for the specific recognition of Rdx.
Only a small number of direct interactions govern mutual recognition of RdxR and
Rdx, corroborating the transient nature of the complex. The shortest distance
between the redox centers is observed to be 6.2 A.
|
|
|
|
|
|
| |
Selected figure(s)
|
|
|
|
| |
|
|

|
|

|
|
Figure 2.
Fig. 2. Structure of the Rdx–RdxR complex. (A) Cartoon
representation. RdxR colors are as in Fig. 1A, and Rdx is in
dark red. FAD, modeled NADH (translucent), and cysteine residues
of the iron-binding site are shown in ball-and-stick mode, and
Fe^3+ is shown as a green sphere. Anomalous-difference electron
density (red) contoured at 6.0  documents the presence
of the Fe^3+. Amino acid exchanges between RubA1 and RubA2 are
indicated as spheres in Rdx (conservative, orange;
nonconservative, blue). (B) Interactions surrounding the
redox-active site of the complex. Interacting residues are shown
as ball-and-stick models (C of RdxR, yellow, and of Rdx,
red-brown). FAD is shown in ball-and-stick mode. (C) Binding
curves for the interaction of RdxR to Fe^3+ (red)- and Ni^2+
(green)-substituted Rdx. documents the presence
of the Fe^3+. Amino acid exchanges between RubA1 and RubA2 are
indicated as spheres in Rdx (conservative, orange;
nonconservative, blue). (B) Interactions surrounding the
redox-active site of the complex. Interacting residues are shown
as ball-and-stick models (C of RdxR, yellow, and of Rdx,
red-brown). FAD is shown in ball-and-stick mode. (C) Binding
curves for the interaction of RdxR to Fe^3+ (red)- and Ni^2+
(green)-substituted Rdx.
|
|
Figure 3.
Fig. 3. Phylogenetic topology of structurally characterized
enzymes functionally and/or structurally related to RdxR. (A)
FNR, plant-type ferredoxin reductase; AdxR, adrenodoxin
reductase; AIF, apoptosis inducing factor; LpdR, lipoamide
reductase; TptR, trypanothione reductase. Sequence identity to
RdxR is indicated by percentage. (B) Superposition of
uncomplexed (gray) and complexed RdxR (colored as in Fig. 1A).
Cofactors are shown as ball-and-stick models. For clarity, the
position of Rdx is indicated by a brown sphere. (C) Comparison
of the Rdx (red) and Trx (black) binding sites of RdxR and TrxR.
For clarity, only the molecular surface of RdxR is shown.  G^0
optimized ET rates (42) between FAD and each point of the RdxR
surface are indicated by a color gradient (red, high ET rate;
green, medium; blue, low). (Inset) Close-up view of the
cofactors involved. G^0
optimized ET rates (42) between FAD and each point of the RdxR
surface are indicated by a color gradient (red, high ET rate;
green, medium; blue, low). (Inset) Close-up view of the
cofactors involved.
|
|
|
|
| |
Figures were
selected
by the author.
|
|
|
|
| |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| |
|
|
|
|
| |
Crude oil spills represent a major ecological threat due to the chemical inertness of the constituent n-alkanes. The Gram-negative bacterium Pseudomonas aeruginosa is one of the few bacterial species able to metabolize such compounds. Three chromosomal genes, rubB, rubA1 and rubA2 coding for an NAD(P)H:rubredoxin oxidoreductase (RdxR) and two rubredoxins (Rdxs) are indispensable for this ability. They constitute an electron transport (ET) pathway that shuttles reducing equivalents from carbon metabolism to the membrane-bound alkane hydroxylases AlkB1 and AlkB2. The RdxR/Rdx system is also crucial as part of the oxidative stress response in archaea or anaerobic bacteria. The redox couple has been analyzed in detail as a model system for ET processes. The crystal structure of RdxR, refined at 2.40 resolution, reveals two domains, a cofactor-binding domains and a C-terminal domain essential for the specific recognition of Rdx. The crystal structure of the RdxR/Rdx complex refined at 2.45Å indicates that mutual recognition is governed by a small number of direct interactions, corroborating the transient nature of the complex. The shortest distance between the redox centers is observed to be 6.2Å.
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
 |
 |
 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Literature references that cite this PDB file's key reference
|
|
|
| |
PubMed id
|
|
Reference
|
|
|
|
|
|
K.Nishikawa,
Y.Shomura,
S.Kawasaki,
Y.Niimura,
and
Y.Higuchi
(2010).
Crystal structure of NADH:rubredoxin oxidoreductase from Clostridium acetobutylicum: a key component of the dioxygen scavenging system in obligatory anaerobes.
|
| |
Proteins,
78,
1066-1070.
|
|
|
PDB code:
|
|
|
|
|
|
|
|
S.Garvis,
A.Munder,
G.Ball,
S.de Bentzmann,
L.Wiehlmann,
J.J.Ewbank,
B.Tümmler,
and
A.Filloux
(2009).
Caenorhabditis elegans semi-automated liquid screen reveals a specialized role for the chemotaxis gene cheB2 in Pseudomonas aeruginosa virulence.
|
| |
PLoS Pathog,
5,
e1000540.
|
|
|
|
|
|
|
E.Bitto,
C.A.Bingman,
L.Bittova,
D.A.Kondrashov,
R.M.Bannen,
B.G.Fox,
J.L.Markley,
and
G.N.Phillips
(2008).
Structure of human J-type co-chaperone HscB reveals a tetracysteine metal-binding domain.
|
| |
J Biol Chem,
283,
30184-30192.
|
|
|
PDB code:
|
|
|
|
|
|
|
|
I.Lüke,
G.Butland,
K.Moore,
G.Buchanan,
V.Lyall,
S.A.Fairhurst,
J.F.Greenblatt,
A.Emili,
T.Palmer,
and
F.Sargent
(2008).
Biosynthesis of the respiratory formate dehydrogenases from Escherichia coli: characterization of the FdhE protein.
|
| |
Arch Microbiol,
190,
685-696.
|
|
|
|
|
|
|
M.Medina,
R.Abagyan,
C.Gómez-Moreno,
and
J.Fernandez-Recio
(2008).
Docking analysis of transient complexes: interaction of ferredoxin-NADP+ reductase with ferredoxin and flavodoxin.
|
| |
Proteins,
72,
848-862.
|
|
|
|
|
|
|
M.V.Petoukhov,
J.B.Vicente,
P.B.Crowley,
M.A.Carrondo,
M.Teixeira,
and
D.I.Svergun
(2008).
Quaternary structure of flavorubredoxin as revealed by synchrotron radiation small-angle X-ray scattering.
|
| |
Structure,
16,
1428-1436.
|
|
|
|
|
|
|
P.B.Crowley,
P.Ganji,
and
H.Ibrahim
(2008).
Protein surface recognition: structural characterisation of cytochrome c-porphyrin complexes.
|
| |
Chembiochem,
9,
1029-1033.
|
|
|
|
|
|
The most recent references are shown first.
Citation data come partly from CiteXplore and partly
from an automated harvesting procedure. Note that this is likely to be
only a partial list as not all journals are covered by
either method. However, we are continually building up the citation data
so more and more references will be included with time.
Where a reference describes a PDB structure, the PDB
code is
shown on the right.
|
 |



 Links
Links














