 |
PDBsum entry 2rj4

|
|
|
|
|
 |
Contents |
 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
* Residue conservation analysis
|
|
|
|
|
|
|
|
|
|
|
Enzyme class 1:
|
|
E.C.2.4.1.37
- fucosylgalactoside 3-alpha-galactosyltransferase.
|
|
|
|
|
|
|
Reaction:
|
|
an alpha-L-fucosyl-(1->2)-beta-D-galactosyl derivative + UDP-alpha-D- galactose = an alpha-D-galactosyl-(1->3)-[alpha-L-fucosyl-(1->2)]-beta-D- galactosyl derivative + UDP + H+
|
|
|
|
|
|
alpha-L-fucosyl-(1->2)-beta-D-galactosyl derivative
|
+
|
UDP-alpha-D- galactose
|
=
|
alpha-D-galactosyl-(1->3)-[alpha-L-fucosyl-(1->2)]-beta-D- galactosyl derivative
|
+
|
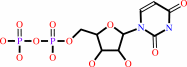 UDP
UDP
|
+
|
H(+)
Bound ligand (Het Group name = )
corresponds exactly
|
|
|
|
|
|
|
|
|
|
Enzyme class 2:
|
|
E.C.2.4.1.40
- glycoprotein-fucosylgalactoside alpha-N-acetylgalactosaminyltransferase.
|
|
|
|
|
|
|
Reaction:
|
|
an alpha-L-fucosyl-(1->2)-beta-D-galactosyl derivative + UDP-N-acetyl- alpha-D-galactosamine = an N-acetyl-alpha-D-galactosaminyl-(1->3)-[alpha- L-fucosyl-(1->2)]-beta-D-galactosyl derivative + UDP + H+
|
|
|
|
|
|
alpha-L-fucosyl-(1->2)-beta-D-galactosyl derivative
|
+
|
UDP-N-acetyl- alpha-D-galactosamine
|
=
|
N-acetyl-alpha-D-galactosaminyl-(1->3)-[alpha- L-fucosyl-(1->2)]-beta-D-galactosyl derivative
|
+
|
UDP
|
+
|
H(+)
Bound ligand (Het Group name = )
corresponds exactly
|
|
|
|
|
|
|
|
|
|
|
|
|
Note, where more than one E.C. class is given (as above), each may
correspond to a different protein domain or, in the case of polyprotein
precursors, to a different mature protein.
|
|
|
|
Molecule diagrams generated from .mol files obtained from the
KEGG ftp site
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
| |
|
|
| |
|
DOI no:
|
J Biol Chem
283:10097-10108
(2008)
|
|
PubMed id:
|
|
|
|
|
|
| |
|
ABO(H) blood group A and B glycosyltransferases recognize substrate via specific conformational changes.
|
|
J.A.Alfaro,
R.B.Zheng,
M.Persson,
J.A.Letts,
R.Polakowski,
Y.Bai,
S.N.Borisova,
N.O.Seto,
T.L.Lowary,
M.M.Palcic,
S.V.Evans.
|
|
|
|
|
| |
ABSTRACT
|
|
|
|
| |
|
|
The final step in the enzymatic synthesis of the ABO(H) blood group A and B
antigens is catalyzed by two closely related glycosyltransferases, an
alpha-(1-->3)-N-acetylgalactosaminyltransferase (GTA) and an
alpha-(1-->3)-galactosyltransferase (GTB). Of their 354 amino acid residues,
GTA and GTB differ by only four "critical" residues. High resolution structures
for GTB and the GTA/GTB chimeric enzymes GTB/G176R and GTB/G176R/G235S bound to
a panel of donor and acceptor analog substrates reveal "open," "semi-closed,"
and "closed" conformations as the enzymes go from the unliganded to the liganded
states. In the open form the internal polypeptide loop (amino acid residues
177-195) adjacent to the active site in the unliganded or H antigen-bound
enzymes is composed of two alpha-helices spanning Arg(180)-Met(186) and
Arg(188)-Asp(194), respectively. The semi-closed and closed forms of the enzymes
are generated by binding of UDP or of UDP and H antigen analogs, respectively,
and show that these helices merge to form a single distorted helical structure
with alternating alpha-3(10)-alpha character that partially occludes the active
site. The closed form is distinguished from the semi-closed form by the ordering
of the final nine C-terminal residues through the formation of hydrogen bonds to
both UDP and H antigen analogs. The semi-closed forms for various mutants
generally show significantly more disorder than the open forms, whereas the
closed forms display little or no disorder depending strongly on the identity of
residue 176. Finally, the use of synthetic analogs reveals how H antigen
acceptor binding can be critical in stabilizing the closed conformation. These
structures demonstrate a delicately balanced substrate recognition mechanism and
give insight on critical aspects of donor and acceptor specificity, on the order
of substrate binding, and on the requirements for catalysis.
|
|
|
|
|
|
| |
Selected figure(s)
|
|
|
|
| |
|
|

|
|

|
|
Figure 1.
FIGURE 1. Conformational changes associated with substrate
binding. a, superimposition of unliganded ABBB in the open form
(white) with AABB+UDP-Gal+DA in the closed form (yellow/red)
showing the internal and C-terminal loops (red), UDP-Gal and DA
(orange), and Mn^2+ (blue), and the location of Arg^176. b,
expanded view about the active site with an arrow indicating the
movement of the internal loop toward the donor in going from the
open state to both the semi-closed or closed states, and showing
the ordering of the C-terminal residues to form the closed
state. c, stereoview of electron density corresponding to the
internal loop in AABB+UDP showing two distinct conformations of
the enzyme (at 50% occupancy) corresponding to the open (yellow)
and semi-closed (green) forms of the enzyme. The disorder
converges at Met^189 (gray). d, the transformation of the
internal loop (residues 176-195) from the open (left) to the
semi-closed (right) conformation is accomplished by the merger
of two  -helices
(Arg^180-Met^186 and Arg^187-Asp^194) into a distorted helical
structure with alternating -3[10]- character. The pivot
point is indicated by a star. -helices
(Arg^180-Met^186 and Arg^187-Asp^194) into a distorted helical
structure with alternating -3[10]- character. The pivot
point is indicated by a star.
|
|
Figure 4.
FIGURE 4. Schematic representation of donor and acceptor
recognition in GTB. The chimeric enzyme AABB displays the closed
form when bound to UDP-Gal and DA, which allows for a complete
characterization of substrate recognition. The acceptor Gal-O-3
is modeled and does not appear in the 3-deoxy acceptor DA.
|
|
|
|
|
|
| |
The above figures are
reprinted
by permission from the ASBMB:
J Biol Chem
(2008,
283,
10097-10108)
copyright 2008.
|
|
| |
Figures were
selected
by the author.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
 |
 |
|
Literature references that cite this PDB file's key reference
|
|
|
| |
PubMed id
|
|
Reference
|
|
|
|
|
|
N.Soya,
Y.Fang,
M.M.Palcic,
and
J.S.Klassen
(2011).
Trapping and characterization of covalent intermediates of mutant retaining glycosyltransferases.
|
| |
Glycobiology,
21,
547-552.
|
|
|
|
|
|
|
B.Schuman,
M.Persson,
R.C.Landry,
R.Polakowski,
J.T.Weadge,
N.O.Seto,
S.N.Borisova,
M.M.Palcic,
and
S.V.Evans
(2010).
Cysteine-to-serine mutants dramatically reorder the active site of human ABO(H) blood group B glycosyltransferase without affecting activity: structural insights into cooperative substrate binding.
|
| |
J Mol Biol,
402,
399-411.
|
|
|
PDB codes:
|
|
|
|
|
|
|
|
C.Rademacher,
J.Landström,
N.Sindhuwinata,
M.M.Palcic,
G.Widmalm,
and
T.Peters
(2010).
NMR-based exploration of the acceptor binding site of human blood group B galactosyltransferase with molecular fragments.
|
| |
Glycoconj J,
27,
349-358.
|
|
|
|
|
|
|
F.Yamamoto,
M.Yamamoto,
and
A.Blancher
(2010).
Generation of histo-blood group B transferase by replacing the N-acetyl-D-galactosamine recognition domain of human A transferase with the galactose-recognition domain of evolutionarily related murine alpha1,3-galactosyltransferase.
|
| |
Transfusion,
50,
622-630.
|
|
|
|
|
|
|
N.Sindhuwinata,
E.Munoz,
F.J.Munoz,
M.M.Palcic,
H.Peters,
and
T.Peters
(2010).
Binding of an acceptor substrate analog enhances the enzymatic activity of human blood group B galactosyltransferase.
|
| |
Glycobiology,
20,
718-723.
|
|
|
|
|
|
|
T.Pesnot,
M.M.Palcic,
and
G.K.Wagner
(2010).
A novel fluorescent probe for retaining galactosyltransferases.
|
| |
Chembiochem,
11,
1392-1398.
|
|
|
|
|
|
|
T.Pesnot,
R.Jørgensen,
M.M.Palcic,
and
G.K.Wagner
(2010).
Structural and mechanistic basis for a new mode of glycosyltransferase inhibition.
|
| |
Nat Chem Biol,
6,
321-323.
|
|
|
PDB codes:
|
|
|
|
|
|
|
|
J.Zschocke
(2008).
Dominant versus recessive: molecular mechanisms in metabolic disease.
|
| |
J Inherit Metab Dis,
31,
599-618.
|
|
|
|
|
|
The most recent references are shown first.
Citation data come partly from CiteXplore and partly
from an automated harvesting procedure. Note that this is likely to be
only a partial list as not all journals are covered by
either method. However, we are continually building up the citation data
so more and more references will be included with time.
Where a reference describes a PDB structure, the PDB
codes are
shown on the right.
|
|



 Links
Links












