 |
PDBsum entry 2jbt

|
|
|
|
|
|

|

|

|

|
|

|
|

|
|

|
|

|
|

|
|

|
|
|
 |

|

|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Oxidoreductase
|
PDB id
|
|
|
|
2jbt
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|

|

|

|
|
|
|
|
|
|
|
 |
Contents |
 |
|
|
|
|
|
|
|
|
|
|
|
|
* Residue conservation analysis
|
|
|
|
|
|
|
|
|
|
|
Enzyme class:
|
|
E.C.1.14.14.9
- 4-hydroxyphenylacetate 3-monooxygenase.
|
|
|
|
|
|
|
Reaction:
|
|
4-hydroxyphenylacetate + FADH2 + O2 = 3,4-dihydroxyphenylacetate + FAD + H2O + H+
|
|
|
|
|
|
 4-hydroxyphenylacetate
4-hydroxyphenylacetate
|
+
|
FADH2
Bound ligand (Het Group name = )
matches with 58.49% similarity
|
+
|
 O2
O2
|
=
|
3,4-dihydroxyphenylacetate
Bound ligand (Het Group name = )
matches with 91.67% similarity
|
+
|
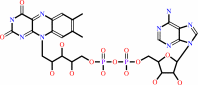 FAD
FAD
|
+
|
H2O
|
+
|
H(+)
|
|
|
|
|
|
|
|
|
|
Cofactor:
|
|
FAD
|
|
|
|
|
|
FAD
|
|
|
|
|
|
|
Molecule diagrams generated from .mol files obtained from the
KEGG ftp site
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
| |
|
|
| |
|
DOI no:
|
Proc Natl Acad Sci U S A
104:1177-1182
(2007)
|
|
PubMed id:
|
|
|
|
|
|
| |
|
Structure of the monooxygenase component of a two-component flavoprotein monooxygenase.
|
|
A.Alfieri,
F.Fersini,
N.Ruangchan,
M.Prongjit,
P.Chaiyen,
A.Mattevi.
|
|
|
|
|
| |
ABSTRACT
|
|
|
|
| |
|
|
p-Hydroxyphenylacetate hydroxylase from Acinetobacter baumannii is a
two-component system consisting of a NADH-dependent FMN reductase and a
monooxygenase (C2) that uses reduced FMN as substrate. The crystal structures of
C2 in the ligand-free and substrate-bound forms reveal a preorganized pocket
that binds reduced FMN without large conformational changes. The Phe-266 side
chain swings out to provide the space for binding p-hydroxyphenylacetate that is
oriented orthogonal to the flavin ring. The geometry of the substrate-binding
site of C2 is significantly different from that of p-hydroxybenzoate
hydroxylase, a single-component flavoenzyme that catalyzes a similar reaction.
The C2 overall structure resembles the folding of medium-chain acyl-CoA
dehydrogenase. An outstanding feature in the C2 structure is a cavity located in
front of reduced FMN; it has a spherical shape with a 1.9-A radius and a 29-A3
volume and is interposed between the flavin C4a atom and the substrate atom to
be hydroxylated. The shape and position of this cavity are perfectly fit for
housing the oxygen atoms of the flavin C4a-hydroperoxide intermediate that is
formed upon reaction of the C2-bound reduced flavin with molecular oxygen. The
side chain of His-396 is predicted to act as a hydrogen-bond donor to the oxygen
atoms of the intermediate. This architecture promotes the nucleophilic attack of
the substrate onto the terminal oxygen of the hydroperoxyflavin. Comparative
analysis with the structures of other flavoenzymes indicates that a distinctive
feature of monooxygenases is the presence of specific cavities that encapsulate
and stabilize the crucial hydroperoxyflavin intermediate.
|
|
|
|
|
|
| |
Selected figure(s)
|
|
|
|
| |
|
|

|
|

|
|
Figure 4.
Fig. 4. Interactions between FMNH^– and C[2]. Distances
are in Å and refer to subunit A of C[2]:FMNH^–:HPA
structure; # indicates residues from subunit D.
|
|
Figure 8.
Fig. 8. Comparison between the active sites of C[2] and
glucose oxidase, a fast oxygen-reacting oxidase that does not
stabilize flavin C4a-hydroperoxide. A histidine side chain
located in front of the flavin is present in both enzymes
(His-516 in glucose oxidase and His-396 in C[2]), although with
shifted position with respect to the flavin C4 locus. The
different positioning of the active site His residues is
visualized by superimposing the flavin ring atoms of glucose
oxidase (PDB ID code 1GAL) on the equivalent atoms of C[2]. The
picture shows His-516 of glucose oxidase (carbons in dark gray)
and the model for the C4a-hydroperoxyflavin and His-396 of C[2]
(carbons in pale pink). Site-directed mutagenesis studies have
shown that the protonated form of His-516 is largely responsible
for the high reaction rate of oxygen with the reduced flavin in
glucose oxidase (31).
|
|
|
|
| |
Figures were
selected
by an automated process.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
 |
 |
|
Literature references that cite this PDB file's key reference
|
|
|
| |
PubMed id
|
|
Reference
|
|
|
|
|
|
D.R.Boyd,
N.D.Sharma,
P.J.Stevenson,
M.Blain,
C.McRoberts,
J.T.Hamilton,
J.M.Argudo,
H.Mundi,
L.A.Kulakov,
and
C.C.Allen
(2011).
Dioxygenase-catalysed cis-dihydroxylation of meta-substituted phenols to yield cyclohexenone cis-diol and derived enantiopure cis-triol metabolites.
|
| |
Org Biomol Chem,
9,
1479-1490.
|
|
|
|
|
|
|
T.Awakawa,
N.Fujita,
M.Hayakawa,
Y.Ohnishi,
and
S.Horinouchi
(2011).
Characterization of the Biosynthesis Gene Cluster for Alkyl-O-Dihydrogeranyl-Methoxyhydroquinones in Actinoplanes missouriensis.
|
| |
Chembiochem,
12,
439-448.
|
|
|
|
|
|
|
M.Morikawa
(2010).
Dioxygen activation responsible for oxidation of aliphatic and aromatic hydrocarbon compounds: current state and variants.
|
| |
Appl Microbiol Biotechnol,
87,
1595-1603.
|
|
|
|
|
|
|
S.Chakraborty,
M.Ortiz-Maldonado,
B.Entsch,
and
D.P.Ballou
(2010).
Studies on the mechanism of p-hydroxyphenylacetate 3-hydroxylase from Pseudomonas aeruginosa: a system composed of a small flavin reductase and a large flavin-dependent oxygenase.
|
| |
Biochemistry,
49,
372-385.
|
|
|
|
|
|
|
U.E.Ukaegbu,
A.Kantz,
M.Beaton,
G.T.Gassner,
and
A.C.Rosenzweig
(2010).
Structure and ligand binding properties of the epoxidase component of styrene monooxygenase .
|
| |
Biochemistry,
49,
1678-1688.
|
|
|
PDB code:
|
|
|
|
|
|
|
|
W.Pitsawong,
J.Sucharitakul,
M.Prongjit,
T.C.Tan,
O.Spadiut,
D.Haltrich,
C.Divne,
and
P.Chaiyen
(2010).
A conserved active-site threonine is important for both sugar and flavin oxidations of pyranose 2-oxidase.
|
| |
J Biol Chem,
285,
9697-9705.
|
|
|
PDB codes:
|
|
|
|
|
|
|
|
R.Baron,
C.Riley,
P.Chenprakhon,
K.Thotsaporn,
R.T.Winter,
A.Alfieri,
F.Forneris,
W.J.van Berkel,
P.Chaiyen,
M.W.Fraaije,
A.Mattevi,
and
J.A.McCammon
(2009).
Multiple pathways guide oxygen diffusion into flavoenzyme active sites.
|
| |
Proc Natl Acad Sci U S A,
106,
10603-10608.
|
|
|
|
|
|
|
J.Valton,
C.Mathevon,
M.Fontecave,
V.Nivière,
and
D.P.Ballou
(2008).
Mechanism and regulation of the Two-component FMN-dependent monooxygenase ActVA-ActVB from Streptomyces coelicolor.
|
| |
J Biol Chem,
283,
10287-10296.
|
|
|
|
|
|
|
N.R.Kwon,
J.C.Chae,
K.Y.Choi,
M.Yoo,
G.J.Zylstra,
Y.M.Kim,
B.S.Kang,
and
E.Kim
(2008).
Identification of functionally important amino acids in a novel indigo-producing oxygenase from Rhodococcus sp. strain T104.
|
| |
Appl Microbiol Biotechnol,
79,
417-422.
|
|
|
|
|
|
|
Y.Hu,
A.Al-Mestarihi,
C.L.Grimes,
D.Kahne,
and
B.O.Bachmann
(2008).
A unifying nitrososynthase involved in nitrosugar biosynthesis.
|
| |
J Am Chem Soc,
130,
15756-15757.
|
|
|
|
|
|
|
P.Nicholls
(2007).
The oxygenase-peroxidase theory of Bach and Chodat and its modern equivalents: change and permanence in scientific thinking as shown by our understanding of the roles of water, peroxide, and oxygen in the functioning of redox enzymes.
|
| |
Biochemistry (Mosc),
72,
1039-1046.
|
|
|
|
|
|
The most recent references are shown first.
Citation data come partly from CiteXplore and partly
from an automated harvesting procedure. Note that this is likely to be
only a partial list as not all journals are covered by
either method. However, we are continually building up the citation data
so more and more references will be included with time.
Where a reference describes a PDB structure, the PDB
code is
shown on the right.
|
|


 Links
Links












