 |
PDBsum entry 2j9f

|
|
|
|
|
|

|

|

|

|
|

|
|

|
|

|
|

|
|

|
|

|
|

|
|

|
|
|
 |

|

|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Oxidoreductase
|
PDB id
|
|
|
|
2j9f
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|

|

|

|
|
|
|
|
|
|
|
 |
Contents |
 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
* Residue conservation analysis
|
|
|
|
|
|
PDB id:
|
|
|
|
| Name: |
|
Oxidoreductase
|
|
|
Title:
|
|
Human branched-chain alpha-ketoacid dehydrogenase-decarboxylase e1b
|
|
Structure:
|
|
2-oxoisovalerate dehydrogenase alpha subunit. Chain: a, c. Synonym: branched-chain alpha-keto acid dehydrogenase e1 component alpha chain, bckdh e1-alpha, bckde1a. Engineered: yes. Mutation: yes. 2-oxoisovalerate dehydrogenase beta subunit. Chain: b, d. Synonym: branched-chain alpha-keto acid dehydrogenase e1 component
|
|
Source:
|
|
Homo sapiens. Human. Organism_taxid: 9606. Expressed in: escherichia coli bl21. Expression_system_taxid: 511693. Expressed in: escherichia coli.
|
|
Resolution:
|
|
|
1.88Å
|
R-factor:
|
0.171
|
R-free:
|
0.200
|
|
|
Authors:
|
|
L.Jun,M.Machius,J.L.Chuang,R.M.Wynn,D.T.Chuang
|
Key ref:

|
|
J.Li
et al.
(2007).
The two active sites in human branched-chain alpha-keto acid dehydrogenase operate independently without an obligatory alternating-site mechanism.
J Biol Chem,
282,
11904-11913.
PubMed id:
DOI:

|
|
|
Date:
|
|
|
07-Nov-06
|
Release date:
|
27-Feb-07
|
|
|
|
|
|
|
PROCHECK
|

|
|
|
|
|
Headers
|
|
|
|
References
|
|
|
|
|
|
|
|
|
|
|
|
Enzyme class:
|
|
Chains A, B, C, D:
E.C.1.2.4.4
- 3-methyl-2-oxobutanoate dehydrogenase (2-methylpropanoyl-transferring).
|
|
|
|
|
|
|
Pathway:
|
|
Oxo-acid dehydrogenase complexes
|
|
|
|
|
|
Reaction:
|
|
N6-[(R)-lipoyl]-L-lysyl-[protein] + 3-methyl-2-oxobutanoate + H+ = N6-[(R)-S(8)-2-methylpropanoyldihydrolipoyl]-L-lysyl-[protein] + CO2
|
|
|
|
|
|
N(6)-[(R)-lipoyl]-L-lysyl-[protein]
|
+
|
3-methyl-2-oxobutanoate
Bound ligand (Het Group name = )
matches with 55.56% similarity
|
+
|
H(+)
|
=
|
N(6)-[(R)-S(8)-2-methylpropanoyldihydrolipoyl]-L-lysyl-[protein]
|
+
|
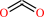 CO2
CO2
|
|
|
|
|
|
|
|
|
|
Cofactor:
|
|
Thiamine diphosphate
|
|
|
|
|
|
Thiamine diphosphate
Bound ligand (Het Group name =
THV)
matches with 83.87% similarity
|
|
|
|
|
|
|
Molecule diagrams generated from .mol files obtained from the
KEGG ftp site
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
| |
|
|
| |
|
DOI no:
|
J Biol Chem
282:11904-11913
(2007)
|
|
PubMed id:
|
|
|
|
|
|
| |
|
The two active sites in human branched-chain alpha-keto acid dehydrogenase operate independently without an obligatory alternating-site mechanism.
|
|
J.Li,
M.Machius,
J.L.Chuang,
R.M.Wynn,
D.T.Chuang.
|
|
|
|
|
| |
ABSTRACT
|
|
|
|
| |
|
|
A long standing controversy is whether an alternating activesite mechanism
occurs during catalysis in thiamine diphosphate (ThDP)-dependent enzymes. We
address this question by investigating the ThDP-dependent
decarboxylase/dehydrogenase (E1b) component of the mitochondrial branched-chain
alpha-keto acid dehydrogenase complex (BCKDC). Our crystal structure reveals
that conformations of the two active sites in the human E1b heterotetramer
harboring the reaction intermediate are identical. Acidic residues in the core
of the E1b heterotetramer, which align with the proton-wire residues proposed to
participate in active-site communication in the related pyruvate dehydrogenase
from Bacillus stearothermophilus, are mutated. Enzyme kinetic data show that,
except in a few cases because of protein misfolding, these alterations are
largely without effect on overall activity of BCKDC, ruling out the requirement
of a proton-relay mechanism in E1b. BCKDC overall activity is nullified at 50%
phosphorylation of E1b, but it is restored to nearly half of the
pre-phosphorylation level after dissociation and reconstitution of BCKDC with
the same phosphorylated E1b. The results suggest that the abolition of overall
activity likely results from the specific geometry of the half-phosphorylated
E1b in the BCKDC assembly and not due to a disruption of the alternating
active-site mechanism. Finally, we show that a mutant E1b containing only one
functional active site exhibits half of the wild-type BCKDC activity, which
directly argues against the obligatory communication between active sites. The
above results provide evidence that the two active sites in the E1b
heterotetramer operate independently during the ThDP-dependent decarboxylation
reaction.
|
|
|
|
|
|
| |
Selected figure(s)
|
|
|
|
| |
|
|

|
|

|
|
Figure 1.
FIGURE 1. Identical conformation in the two active sites of
the human E1b heterotetramer. Ribbon representation of S320P
 human E1b in the
crystal form containing an entire heterotetramer in the
asymmetric unit. The N and C termini are indicated. All figures
of molecular structures were created with the program PyMol
(DeLano Scientific, San Carlos, CA). human E1b in the
crystal form containing an entire heterotetramer in the
asymmetric unit. The N and C termini are indicated. All figures
of molecular structures were created with the program PyMol
(DeLano Scientific, San Carlos, CA).
|
|
Figure 4.
FIGURE 4. SDS-PAGE and Western blotting analysis of
affinity-column fractions containing hybrid (S292Q /WT)
E1b. The hybrid E1b contains the S292Q mutation in the
His[6]-tagged subunit, and the
wild-type sequence in the Strep-tagged subunit was prepared,
as described under "Experimental Procedures." A, SDS-PAGE of
peak fractions from a HisTrap column; B, SDS-PAGE of peak
fractions from a consecutive Strep-Tactin Superflow column; C,
Western blotting with probes for the His[6] or Strep tag. Hyb,
hybrid E1b.
|
|
|
|
|
|
| |
The above figures are
reprinted
by permission from the ASBMB:
J Biol Chem
(2007,
282,
11904-11913)
copyright 2007.
|
|
| |
Figures were
selected
by an automated process.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
 |
 |
|
Literature references that cite this PDB file's key reference
|
|
|
| |
PubMed id
|
|
Reference
|
|
|
|
|
|
S.Kale,
and
F.Jordan
(2009).
Conformational ensemble modulates cooperativity in the rate-determining catalytic step in the E1 component of the Escherichia coli pyruvate dehydrogenase multienzyme complex.
|
| |
J Biol Chem,
284,
33122-33129.
|
|
|
|
|
|
|
M.Kato,
R.M.Wynn,
J.L.Chuang,
S.C.Tso,
M.Machius,
J.Li,
and
D.T.Chuang
(2008).
Structural basis for inactivation of the human pyruvate dehydrogenase complex by phosphorylation: role of disordered phosphorylation loops.
|
| |
Structure,
16,
1849-1859.
|
|
|
PDB codes:
|
|
|
|
|
|
|
|
R.M.Wynn,
M.Kato,
J.L.Chuang,
S.C.Tso,
J.Li,
and
D.T.Chuang
(2008).
Pyruvate dehydrogenase kinase-4 structures reveal a metastable open conformation fostering robust core-free basal activity.
|
| |
J Biol Chem,
283,
25305-25315.
|
|
|
PDB codes:
|
|
|
|
|
|
|
|
X.Y.Pei,
C.M.Titman,
R.A.Frank,
F.J.Leeper,
and
B.F.Luisi
(2008).
Snapshots of catalysis in the E1 subunit of the pyruvate dehydrogenase multienzyme complex.
|
| |
Structure,
16,
1860-1872.
|
|
|
PDB codes:
|
|
|
|
|
|
|
The most recent references are shown first.
Citation data come partly from CiteXplore and partly
from an automated harvesting procedure. Note that this is likely to be
only a partial list as not all journals are covered by
either method. However, we are continually building up the citation data
so more and more references will be included with time.
Where a reference describes a PDB structure, the PDB
codes are
shown on the right.
|
|
|




 Links
Links













