 |
PDBsum entry 2e3c

|
|
|
|
|
 |
Contents |
 |
|
|
|
|
|
|
|
|
|
|
* Residue conservation analysis
|
|
|
|
|
|
|
|
|
|
|
Enzyme class:
|
|
E.C.6.1.1.26
- pyrrolysine--tRNA(Pyl) ligase.
|
|
|
|
|
|
|
Reaction:
|
|
tRNA(Pyl) + L-pyrrolysine + ATP = L-pyrrolysyl-tRNA(Pyl) + AMP + diphosphate
|
|
|
|
|
|
 tRNA(Pyl)
tRNA(Pyl)
|
+
|
 L-pyrrolysine
L-pyrrolysine
|
+
|
 ATP
ATP
|
=
|
 L-pyrrolysyl-tRNA(Pyl)
L-pyrrolysyl-tRNA(Pyl)
|
+
|
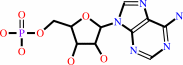 AMP
AMP
|
+
|
 diphosphate
diphosphate
|
|
|
|
|
|
|
|
|
|
|
|
|
Molecule diagrams generated from .mol files obtained from the
KEGG ftp site
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
| |
|
|
| |
|
DOI no:
|
J Mol Biol
378:634-652
(2008)
|
|
PubMed id:
|
|
|
|
|
|
| |
|
Crystallographic studies on multiple conformational states of active-site loops in pyrrolysyl-tRNA synthetase.
|
|
T.Yanagisawa,
R.Ishii,
R.Fukunaga,
T.Kobayashi,
K.Sakamoto,
S.Yokoyama.
|
|
|
|
|
| |
ABSTRACT
|
|
|
|
| |
|
|
Pyrrolysine, a lysine derivative with a bulky pyrroline ring, is the "22nd"
genetically encoded amino acid. In the present study, the carboxy-terminal
catalytic fragment of Methanosarcina mazei pyrrolysyl-tRNA synthetase (PylRS)
was analyzed by X-ray crystallography and site-directed mutagenesis. The
catalytic fragment ligated tRNA(Pyl) with pyrrolysine nearly as efficiently as
the full-length PylRS. We determined the crystal structures of the PylRS
catalytic fragment in the substrate-free, ATP analogue (AMPPNP)-bound, and
AMPPNP/pyrrolysine-bound forms, and compared them with the previously-reported
PylRS structures. The ordering loop and the motif-2 loop undergo conformational
changes from the "open" states to the "closed" states upon AMPPNP binding. On
the other hand, the beta 7-beta 8 hairpin exhibits multiple conformational
states, the open, intermediate (beta 7-open/beta 8-open and beta 7-closed/beta
8-open), and closed states, which are not induced upon substrate binding. The
PylRS structures with a docked tRNA suggest that the active-site pocket can
accommodate the CCA terminus of tRNA when the motif-2 loop is in the closed
state and the beta 7-beta 8 hairpin is in the open or intermediate state. The
entrance of the active-site pocket is nearly closed in the closed state of the
beta 7-beta 8 hairpin, which may protect the pyrrolysyladenylate intermediate in
the absence of tRNA(Pyl). Moreover, a structure-based mutational analysis
revealed that hydrophobic residues in the amino acid-binding tunnel are
important for accommodating the pyrrolysine side chain and that Asn346 is
essential for anchoring the side-chain carbonyl and alpha-amino groups of
pyrrolysine. In addition, a docking model of PylRS with tRNA was constructed
based on the aspartyl-tRNA synthetase/tRNA structure, and was confirmed by a
mutational analysis.
|
|
|
|
|
|
| |
Selected figure(s)
|
|
|
|
| |
|
|

|
|

|
|
Figure 5.
Fig. 5. Comparison of the β7-β8 hairpin conformations. (a)
A stereo view of superpositioned C^α traces of the
pyrrolysyladenylate-bound (black), the pyrrolysine/AMPPNP-bound
(green), AMPPNP-bound (yellow), and apo forms (red). The
locations of the C^α atoms for residues 377–387 are numbered,
with the same coloring as in each trace. (b and c) Stereo views
of the superposition of the pyrrolysyladenylate-bound (gray),
pyrrolysine/AMPPNP-bound (green), AMPPNP-bound (yellow), and apo
forms (vermilion). Val377, Gly378, and Asp379 (b), and Tyr384,
Gly385, Asp386, and Thr387 (c) are represented by ball-and-stick
models. d and e, A stereo view of the 2F[o] – F[c] electron
density map (contoured at 1.1 σ) around the β7-β8 regions.
(d) The AMPPNP-bound form. Direct hydrogen-bonding interactions
of Val402 with Asp386, and of Asp379 with Arg356, and the
water-mediated hydrogen-bonding interaction of Asp379 with
Thr387 are shown by pink, green, and orange dotted lines,
respectively. (e) The AMPPNP/pyrrolysine-bound form. Direct
hydrogen-bonding interactions of Val402 with Asp386, and of
Asp379 with Thr387 are shown by pink and orange dotted lines,
respectively. Gly385, Asp386, and Thr387 in each PylRS(c270)
form adopt the β8-open conformation, as shown in sky blue
circles. On the other hand, Gly378 and Asp379 adopt the β7-open
conformation in the AMPPNP-bound form and the β7-closed
conformation in the pyrrolysine/AMPPNP-bound form, as shown in
purple circles, respectively.
|
|
Figure 6.
Fig. 6. Pyrrolysine recognition by PylRS(c270). (a) The 2F[o]
– F[c] omit map (contoured at 3 σ) of the active site of
PylRS(c270) complexed with pyrrolysine and AMPPNP. The
pyrrolysine molecule was omitted for the map calculation. (b)
Ball-and-stick models of pyrrolysine and AMPPNP bound within the
PylRS(c270) active site. PylRS(c270) is represented as a surface
model, calculated by MSMS.^53 Hydrophobic residues in the
catalytic cleft (Ala302, Leu305, Tyr306, Leu309, Val401, and
Trp417) and Asn346, which hydrogen bonds with pyrrolysine, are
colored yellow and green, respectively. (c) Active-site
mutations of PylRS(c270) affect the aminoacylation activity for
pyrrolysine. The aminoacylation assay conditions are described
in Materials and Methods. A concentration of 9 μM of was used
for the wild-type and mutant PylRS(c270) proteins. Starting from
the left, each lane shows a reaction with the following: no
enzyme; wild-type PylRS(c270); W417A; Y306A; N346A; C348A;
S399A; V401A; L305A; control tRNA^Pyl. d and e, The active sites
of the pyrrolysine/AMPPNP-bound (d) and
pyrrolysyladenylate-bound forms of PylRS(c270) (e).^15 The
β7-β8 hairpin is shown in yellow. The pyrrolysine, AMPPNP,
pyrrolysyladenylate, Arg330, Asn346, and Tyr384 are shown as
ball-and-stick models. Water molecules (Wat) are shown as pink
balls. The class-II aaRS specific motifs, motif 1, motif 2, and
motif 3 (residues 258–268, 329–345, and 418–431,
respectively) are colored red, green, and blue, respectively.
The three hydrogen-bonding interactions of the PylRS(c270)
active site with pyrrolysine or pyrrolysyladenylate in both
forms, and the two additional hydrogen-bonding interactions in
the pyrrolysyladenylate-bound form are represented by black and
red dotted lines, respectively. The interaction between the
pyrrolysine α-carboxyl group and the α-phosphate atom in the
pyrrolysine/AMPPNP-bound form is also shown (pale blue dotted
line).
|
|
|
|
|
|
| |
The above figures are
reprinted
by permission from Elsevier:
J Mol Biol
(2008,
378,
634-652)
copyright 2008.
|
|
| |
Figures were
selected
by an automated process.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
 |
 |
|
Literature references that cite this PDB file's key reference
|
|
|
| |
PubMed id
|
|
Reference
|
|
|
|
|
|
J.Yuan,
P.O'Donoghue,
A.Ambrogelly,
S.Gundllapalli,
R.Lynn Sherrer,
S.Palioura,
M.Simonović,
and
D.Söll
(2010).
Distinct genetic code expansion strategies for selenocysteine and pyrrolysine are reflected in different aminoacyl-tRNA formation systems.
|
| |
FEBS Lett,
584,
342-349.
|
|
|
|
|
|
|
M.Rother,
and
J.A.Krzycki
(2010).
Selenocysteine, pyrrolysine, and the unique energy metabolism of methanogenic archaea.
|
| |
Archaea,
2010,
0.
|
|
|
|
|
|
|
X.Li,
T.Fekner,
and
M.K.Chan
(2010).
N6-(2-(R)-propargylglycyl)lysine as a clickable pyrrolysine mimic.
|
| |
Chem Asian J,
5,
1765-1769.
|
|
|
|
|
|
|
K.Nozawa,
P.O'Donoghue,
S.Gundllapalli,
Y.Araiso,
R.Ishitani,
T.Umehara,
D.Söll,
and
O.Nureki
(2009).
Pyrrolysyl-tRNA synthetase-tRNA(Pyl) structure reveals the molecular basis of orthogonality.
|
| |
Nature,
457,
1163-1167.
|
|
|
PDB codes:
|
|
|
|
|
|
|
|
M.M.Lee,
R.Jiang,
R.Jain,
R.C.Larue,
J.Krzycki,
and
M.K.Chan
(2008).
Structure of Desulfitobacterium hafniense PylSc, a pyrrolysyl-tRNA synthetase.
|
| |
Biochem Biophys Res Commun,
374,
470-474.
|
|
|
PDB code:
|
|
|
|
|
|
|
|
S.Gundllapalli,
A.Ambrogelly,
T.Umehara,
D.Li,
C.Polycarpo,
and
D.Söll
(2008).
Misacylation of pyrrolysine tRNA in vitro and in vivo.
|
| |
FEBS Lett,
582,
3353-3358.
|
|
|
|
|
|
|
T.Yanagisawa,
R.Ishii,
R.Fukunaga,
T.Kobayashi,
K.Sakamoto,
and
S.Yokoyama
(2008).
Multistep engineering of pyrrolysyl-tRNA synthetase to genetically encode N(epsilon)-(o-azidobenzyloxycarbonyl) lysine for site-specific protein modification.
|
| |
Chem Biol,
15,
1187-1197.
|
|
|
PDB codes:
|
|
|
|
|
|
|
The most recent references are shown first.
Citation data come partly from CiteXplore and partly
from an automated harvesting procedure. Note that this is likely to be
only a partial list as not all journals are covered by
either method. However, we are continually building up the citation data
so more and more references will be included with time.
Where a reference describes a PDB structure, the PDB
codes are
shown on the right.
|
|


 Links
Links












