
|
|
|
|
|
 |
Contents |
 |
|
|
|
|
|
|
|
|
|
|
|
|
* Residue conservation analysis
|
|
|
|
|
|
|
|
|
|
Enzyme class 2:
|
|
Chains A, C:
E.C.1.11.1.5
- cytochrome-c peroxidase.
|
|
|
|
|
|
|
Reaction:
|
|
2 Fe(II)-[cytochrome c] + H2O2 + 2 H+ = 2 Fe(III)-[cytochrome c] + 2 H2O
|
|
|
|
|
|
2
×
Fe(II)-[cytochrome c]
|
+
|
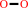 H2O2
H2O2
|
+
|
2
×
H(+)
|
=
|
2
×
Fe(III)-[cytochrome c]
|
+
|
2
×
H2O
|
|
|
|
|
|
|
|
|
|
Cofactor:
|
|
Heme
|
|
|
|
|
|
Heme
Bound ligand (Het Group name =
ZNH)
matches with 95.45% similarity
|
|
|
|
Enzyme class 3:
|
|
Chains B, D:
E.C.?
|
|
|
|
|
|
|
|
|
|
Note, where more than one E.C. class is given (as above), each may
correspond to a different protein domain or, in the case of polyprotein
precursors, to a different mature protein.
|
|
|
|
Molecule diagrams generated from .mol files obtained from the
KEGG ftp site
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
| |
|
|
| |
|
DOI no:
|
Proc Natl Acad Sci U S A
102:15465-15470
(2005)
|
|
PubMed id:
|
|
|
|
|
|
| |
|
Effects of interface mutations on association modes and electron-transfer rates between proteins.
|
|
S.A.Kang,
B.R.Crane.
|
|
|
|
|
| |
ABSTRACT
|
|
|
|
| |
|
|
Although bonding networks determine electron-transfer (ET) rates within
proteins, the mechanism by which structure and dynamics influence ET across
protein interfaces is not well understood. Measurements of photochemically
induced ET and subsequent charge recombination between Zn-porphyrin-substituted
cytochrome c peroxidase and cytochrome c in single crystals correlate reactivity
with defined structures for different association modes of the redox partners.
Structures and ET rates in crystals are consistent with tryptophan oxidation
mediating charge recombination reactions. Conservative mutations at the
interface can drastically affect how the proteins orient and dispose redox
centers. Whereas some configurations are ET inactive, the wild-type complex
exhibits the fastest recombination rate. Other association modes generate ET
rates that do not correlate with predictions based on cofactor separations or
simple bonding pathways. Inhibition of photoinduced ET at <273 K indicates
gating by small-amplitude dynamics, even within the crystal. Thus, different
associations achieve states of similar reactivity, and within those states
conformational fluctuations enable interprotein ET.
|
|
|
|
|
|
| |
Selected figure(s)
|
|
|
|
| |
|
|

|
|

|
|
Figure 1.
Fig. 1. Reactions initiated by photoinduced ET between
ZnCcP and Fe(III) yCc.
|
|
Figure 2.
Fig. 2. Complexes of ZnCcP with various Cc mutants have
different association modes. (A) Representative ribbon diagrams
for ZnCcP bound to Cc variants. From left to right: yCc WT,
F82W, and F82S mutants; yCc F82Y and F82I mutants; horse Cc; yCc
K72S/F82Y. (B) Relative positioning and interface structures for
the ZnCcP:F82W yCc (Left) and ZnCcP:yCc F82I (Right) complexes.
Cc (gray ribbons, top) rotates 90° and
translates8Åinthe F82I mutant generating altered heme
orientations (yellow bonds), interprotein contacts (black and
orange side chains), and intervening solvent structure (red
spheres) with CcP (yellow, below) compared with the F82W
complex. (C) Superposition of ZnCcP from the two ZnCcP: F82S yCc
complexes (red and blue) indicate variability in Cc positioning
within the asymmetric unit.
|
|
|
|
| |
Figures were
selected
by an automated process.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
 |
 |
|
Literature references that cite this PDB file's key reference
|
|
|
| |
PubMed id
|
|
Reference
|
|
|
|
|
|
A.M.Blanco-Rodríguez,
A.J.Di Bilio,
C.Shih,
A.K.Museth,
I.P.Clark,
M.Towrie,
A.Cannizzo,
J.Sudhamsu,
B.R.Crane,
J.Sýkora,
J.R.Winkler,
H.B.Gray,
S.Záliš,
and
A.Vlček
(2011).
Phototriggering electron flow through Re(I)-modified Pseudomonas aeruginosa azurins.
|
| |
Chemistry,
17,
5350-5361.
|
|
|
|
|
|
|
J.Karanicolas,
J.E.Corn,
I.Chen,
L.A.Joachimiak,
O.Dym,
S.H.Peck,
S.Albeck,
T.Unger,
W.Hu,
G.Liu,
S.Delbecq,
G.T.Montelione,
C.P.Spiegel,
D.R.Liu,
and
D.Baker
(2011).
A de novo protein binding pair by computational design and directed evolution.
|
| |
Mol Cell,
42,
250-260.
|
|
|
PDB codes:
|
|
|
|
|
|
|
|
H.K.Ly,
M.A.Marti,
D.F.Martin,
D.Alvarez-Paggi,
W.Meister,
A.Kranich,
I.M.Weidinger,
P.Hildebrandt,
and
D.H.Murgida
(2010).
Thermal fluctuations determine the electron-transfer rates of cytochrome c in electrostatic and covalent complexes.
|
| |
Chemphyschem,
11,
1225-1235.
|
|
|
|
|
|
|
A.M.Carver,
M.De,
H.Bayraktar,
S.Rana,
V.M.Rotello,
and
M.J.Knapp
(2009).
Intermolecular electron-transfer catalyzed on nanoparticle surfaces.
|
| |
J Am Chem Soc,
131,
3798-3799.
|
|
|
|
|
|
|
A.D.Patel,
J.M.Nocek,
and
B.M.Hoffman
(2008).
Kinetic-dynamic model for conformational control of an electron transfer photocycle: mixed-metal hemoglobin hybrids.
|
| |
J Phys Chem B,
112,
11827-11837.
|
|
|
|
|
|
|
Y.C.Kim,
C.Tang,
G.M.Clore,
and
G.Hummer
(2008).
Replica exchange simulations of transient encounter complexes in protein-protein association.
|
| |
Proc Natl Acad Sci U S A,
105,
12855-12860.
|
|
|
|
|
|
|
Y.C.Kim,
and
G.Hummer
(2008).
Coarse-grained models for simulations of multiprotein complexes: application to ubiquitin binding.
|
| |
J Mol Biol,
375,
1416-1433.
|
|
|
|
|
|
|
M.M.Haque,
K.Panda,
J.Tejero,
K.S.Aulak,
M.A.Fadlalla,
A.T.Mustovich,
and
D.J.Stuehr
(2007).
A connecting hinge represses the activity of endothelial nitric oxide synthase.
|
| |
Proc Natl Acad Sci U S A,
104,
9254-9259.
|
|
|
|
|
|
|
A.N.Volkov,
J.A.Worrall,
E.Holtzmann,
and
M.Ubbink
(2006).
Solution structure and dynamics of the complex between cytochrome c and cytochrome c peroxidase determined by paramagnetic NMR.
|
| |
Proc Natl Acad Sci U S A,
103,
18945-18950.
|
|
|
PDB code:
|
|
|
|
|
|
|
|
N.A.Belikova,
Y.A.Vladimirov,
A.N.Osipov,
A.A.Kapralov,
V.A.Tyurin,
M.V.Potapovich,
L.V.Basova,
J.Peterson,
I.V.Kurnikov,
and
V.E.Kagan
(2006).
Peroxidase activity and structural transitions of cytochrome c bound to cardiolipin-containing membranes.
|
| |
Biochemistry,
45,
4998-5009.
|
|
|
|
|
|
The most recent references are shown first.
Citation data come partly from CiteXplore and partly
from an automated harvesting procedure. Note that this is likely to be
only a partial list as not all journals are covered by
either method. However, we are continually building up the citation data
so more and more references will be included with time.
Where a reference describes a PDB structure, the PDB
codes are
shown on the right.
|
 |


 Links
Links














