 |
PDBsum entry 2z6j

|
|
|
|
|
|

|

|

|

|
|

|
|

|
|

|
|

|
|

|
|

|
|

|
|
|
 |

|

|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Oxidoreductase
|
PDB id
|
|
|
|
2z6j
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|

|

|

|
|
|
|
|
|
|
|
 |
Contents |
 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
* Residue conservation analysis
|
|
|
|
|
|
|
|
|
|
|
Enzyme class 1:
|
|
E.C.1.13.12.16
- nitronate monooxygenase.
|
|
|
|
|
|
|
Reaction:
|
|
ethylnitronate + O2 = chemical entity + acetaldehyde + nitrite + H+
|
|
|
|
|
|
 ethylnitronate
ethylnitronate
|
+
|
 O2
O2
|
=
|
chemical entity
|
+
|
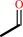 acetaldehyde
acetaldehyde
|
+
|
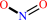 nitrite
nitrite
|
+
|
H(+)
|
|
|
|
|
|
|
|
|
|
Cofactor:
|
|
FMN
|
|
|
|
|
|
FMN
Bound ligand (Het Group name =
FMN)
corresponds exactly
|
|
|
|
Enzyme class 2:
|
|
E.C.1.3.-.-
|
|
|
|
|
|
|
|
|
|
Note, where more than one E.C. class is given (as above), each may
correspond to a different protein domain or, in the case of polyprotein
precursors, to a different mature protein.
|
|
|
|
Molecule diagrams generated from .mol files obtained from the
KEGG ftp site
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
| |
|
|
| |
|
DOI no:
|
Protein Sci
17:691-699
(2008)
|
|
PubMed id:
|
|
|
|
|
|
| |
|
Crystal structure of enoyl-acyl carrier protein reductase (FabK) from Streptococcus pneumoniae reveals the binding mode of an inhibitor.
|
|
J.Saito,
M.Yamada,
T.Watanabe,
M.Iida,
H.Kitagawa,
S.Takahata,
T.Ozawa,
Y.Takeuchi,
F.Ohsawa.
|
|
|
|
|
| |
ABSTRACT
|
|
|
|
| |
|
|
Enoyl-acyl carrier protein (ACP) reductases are critical for bacterial type II
fatty acid biosynthesis and thus are attractive targets for developing novel
antibiotics. We determined the crystal structure of enoyl-ACP reductase (FabK)
from Streptococcus pneumoniae at 1.7 A resolution. There was one dimer per
asymmetric unit. Each subunit formed a triose phosphate isomerase (TIM) barrel
structure, and flavin mononucleotide (FMN) was bound as a cofactor in the active
site. The overall structure was similar to the enoyl-ACP reductase (ER) of
fungal fatty acid synthase and to 2-nitropropane dioxygenase (2-ND) from
Pseudomonas aeruginosa, although there were some differences among these
structures. We determined the crystal structure of FabK in complex with a
phenylimidazole derivative inhibitor to envision the binding site interactions.
The crystal structure reveals that the inhibitor binds to a hydrophobic pocket
in the active site of FabK, and this is accompanied by induced-fit movements of
two loop regions. The thiazole ring and part of the ureido moiety of the
inhibitor are involved in a face-to-face pi-pi stacking interaction with the
isoalloxazine ring of FMN. The side-chain conformation of the proposed catalytic
residue, His144, changes upon complex formation. Lineweaver-Burk plots indicate
that the inhibitor binds competitively with respect to NADH, and uncompetitively
with respect to crotonoyl coenzyme A. We propose that the primary basis of the
inhibitory activity is competition with NADH for binding to FabK, which is the
first step of the two-step ping-pong catalytic mechanism.
|
|
|
|
|
|
| |
Selected figure(s)
|
|
|
|
| |
|
|

|
|

|
|
Figure 1.
Chemical structures of FabK inhibitors, AG205 and compound 1.
Dashed lines enclose similar components of the two compounds.
|
|
Figure 6.
(A) Superposition of the active sites of S. pneumoniae FabK
--compound 1 complex (green) with fungal ER --NADP^+ complex
(cyan). FMN, compound 1, NADP^+, and side chains of the proposed
catalytic histidine residues are shown as stick models.
Non-carbon atoms are colored according to atom type. (B)
Superposition of the active sites of S. pneumoniae FabK
--compound 1 complex (green) with P. aeruginosa 2-ND in complex
with the substrate, 2-nitropropane (pink). (C) Two-step
ping-pong catalytic mechanism of FabK. FabK --FMN represents the
oxidized form of the enzyme, and FabK --FMNH[2] ^[minus sign]
represents the reduced form. R represents the
CH[3](CH[2])[n][minus sign] (n = 1 [minus sign] 12). Compound 1
is considered to be a competitive inhibitor for the first step
of the reaction.
|
|
|
|
|
|
| |
The above figures are
reprinted
from an Open Access publication published by the Protein Society:
Protein Sci
(2008,
17,
691-699)
copyright 2008.
|
|
| |
Figures were
selected
by an automated process.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
 |
 |
|
Literature references that cite this PDB file's key reference
|
|
|
| |
PubMed id
|
|
Reference
|
|
|
|
|
|
L.Zhu,
J.Lin,
J.Ma,
J.E.Cronan,
and
H.Wang
(2010).
Triclosan resistance of Pseudomonas aeruginosa PAO1 is due to FabV, a triclosan-resistant enoyl-acyl carrier protein reductase.
|
| |
Antimicrob Agents Chemother,
54,
689-698.
|
|
|
|
|
|
|
T.Maier,
M.Leibundgut,
D.Boehringer,
and
N.Ban
(2010).
Structure and function of eukaryotic fatty acid synthases.
|
| |
Q Rev Biophys,
43,
373-422.
|
|
|
|
|
|
|
R.P.Massengo-Tiassé,
and
J.E.Cronan
(2009).
Diversity in enoyl-acyl carrier protein reductases.
|
| |
Cell Mol Life Sci,
66,
1507-1517.
|
|
|
|
|
|
|
M.Leibundgut,
T.Maier,
S.Jenni,
and
N.Ban
(2008).
The multienzyme architecture of eukaryotic fatty acid synthases.
|
| |
Curr Opin Struct Biol,
18,
714-725.
|
|
|
|
|
|
|
T.Maier,
M.Leibundgut,
and
N.Ban
(2008).
The crystal structure of a mammalian fatty acid synthase.
|
| |
Science,
321,
1315-1322.
|
|
|
PDB codes:
|
|
|
|
|
|
|
The most recent references are shown first.
Citation data come partly from CiteXplore and partly
from an automated harvesting procedure. Note that this is likely to be
only a partial list as not all journals are covered by
either method. However, we are continually building up the citation data
so more and more references will be included with time.
Where a reference describes a PDB structure, the PDB
codes are
shown on the right.
|
|



 Links
Links












